мембранный потенциал в чем измеряется мв
2_1 Мембранный потенциал покоя кратко (доцент Сазонов В.Ф.)
Введение
Зачем нам нужно знать, что такое потенциал покоя?
Что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку»?
— На эти вопросы мы сможем ответить, если узнаем, как клетка за счёт перераспределения электрических зарядов создаёт себе электрический потенциал на мембране.
Как работает нервная система? С чего в ней всё начинается? Откуда в ней берётся электричество для нервных импульсов?
— На эти вопросы мы также сможем ответить, если узнаем, как нервная клетка создаёт себе электрический потенциал на мембране.
А в основе работы нейрона с нервными импульсами лежит перераспределение электрических зарядов на его мембране и изменение величины электрических потенциалов. Но чтобы потенциал изменять, его нужно для начала иметь. Поэтому можно сказать, что нейрон, готовясь к cвоей нервной работе, создаёт на своей мембране электрический потенциал, как возможность для такой работы.
Определение
В норме, когда клетка готова к работе, у неё уже есть электрический заряд на поверхности мембраны. Он называется мембранный потенциал покоя.
Таким образом, имея электрический потенциал, мембрана может перемещать в клетку или из клетки указанные выше заряженные ионы.
Сущность
1. Поведение мембраны
В поведении мембраны для потенциала покоя важны три процесса:
1) Обмен внутренних ионов натрия на наружные ионы калия. Обменом занимаются специальные транспортные структуры мембраны: ионные насосы-обменники. Таким способом мембрана перенасыщает клетку калием, но обедняет натрием.
2) Открытые калиевые ионные каналы. Через них калий может как заходить в клетку, так и выходить из неё. Он выходит в основном.
2. Поведение ионов калия и натрия
Ионы калия и натрия по-разному перемещаются через мембрану:
2) Через постоянно открытые калиевые каналы калий выходит из клетки, но может и возвращаться в неё обратно через них же.
3) Натрий «хочет» войти в клетку, но «не может», т.к. каналы для него закрыты.
3. Соотношение химической и электрической силы
1) Химическая сила выталкивает калий из клетки, но стремится затянуть в неё натрий.
2) Электрическая сила стремится затянуть в клетку положительно заряженные ионы (как натрий, так и калий).
Формирование потенциала покоя
Натрий-калиевый насос
А всё дело заключается в том, что в мембране нервной клетки постоянно работают насосы-обменники, образованные специальными белками, встроенными в мембрану. Что они делают? Они меняют «собственный» натрий клетки на наружный «чужой» калий. Из-за этого в клетке оказывается в конце концов недостаток натрия, который ушёл на обмен. И в то же время клетка переполняется ионами калия, который в неё натащили эти молекулярные насосы.
Чтобы легче было запомнить, образно можно сказать так: «Клетка любит калий!» (Хотя об истинной любви здесь не может идти и речи!) Поэтому она и затаскивает калий в себя, несмотря на то, что его и так полно. Поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. Поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. Вот что делает любовь, пусть даже не настоящая!
Образно говоря, можно выразиться так:
Создавая потенциал покоя, клетка «заряжается любовью».
Это любовь к двум вещам:
1) любовь клетки к калию,
2) любовь калия к свободе.
Итак, результат деятельности мембранных ионных насосов-обменников таков:
Натрий-калиевый ионный насос-обменник создаёт три потенциала (возможности):
1. Дефицит натрия (Na + ) в клетке.
2. Избыток калия (K + ) в клетке.
Можно сказать так: ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Именно из-за получившегося дефицита натрия в клетку теперь «полезет» этот самый натрий снаружи. Так всегда ведут себя вещества: они стремятся выравнять свою концентрацию во всём объёме раствора.
И в то же время в клетке получился избыток ионов калия по сравнению с наружной средой. Потому что насосы мембраны накачали его в клетку. И он стремится уравнять свою концентрацию внутри и снаружи, и поэтому стремится выйти из клетки.
По закону выравнивания концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку. Но не может, так как мембрана в обычном состоянии плохо его пропускает. Его заходит немножко и клетка его опять тут же обменивает на наружный калий. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Так вот он и выходит наружу через особые белковые дырочки в мембране (ионные каналы).
Анализ
Калий заряжен положительным зарядом, и поэтому он, когда выходит из клетки, выносит из неё не только себя, но и «плюсики» (положительные заряды). На их месте в клетке остаются «минусы» (отрицательные заряды). Это и есть мембранный потенциал покоя!
Заключение

Рис. Схема формирования потенциала покоя (ПП). Автор благодарит Попову Екатерину Юрьевну за помощь в создании рисунка.
Составные части потенциала покоя
Обратите внимание на то, что для создания потенциала покоя нужны затраты энергии. Эти затраты производятся ионными насосами, которые обменивают «свой» внутренний натрий (ионы Na + ) на «чужой» внешний калий (K + ). Вспомним, что ионные насосы являются ферментами АТФазами и расщепляют АТФ, получая из неё энергию на указанный обмен ионов разного типа друг на друга.Тут очень важно понять, что с мембраной «работают» сразу 2 потенциала: химический (концентрационный градиент ионов) и электрический (разность электрических потенциалов по разные стороны мембраны). Ионы перемещаются в ту или иную сторону под действием обеих этих сил, на которые и тратится энергия. При этом один из двух потенциалов (химический или электрический) уменьшается, а другой увеличивается. Разумеется, если рассматривать электрический потенциал (разность потенциалов) отдельно, то не будут учитываться «химические» силы, перемещающие ионы. И тогда может сложиться неверное впечатление о том, что энергия на движение ионо берётся как бы ниоткуда. Но это не так. Необходимо рассматривать обе силы: химическую и электрическую. При этом крупные молекулы с отрицательными зарядами, находящиеся внутри клетки играют роль «статистов», т.к. их не перемещают через мембрану ни химические, ни электрические силы. Поэтому эти отрицательные частицы обычно и не рассматривают, хотя они существуют и именно они обеспечивают отрицательную сторону разности потенциалов между внутренней и наружной сторонами мембраны. А вот шустрые ионы калия, как раз способны к перемещению, и именно их утечка из клетки под действием химических сил создаёт львиную долю электрического потенциала (разности потенциалов). Ведь именно ионы калия перемещают на наружную сторону мембраны положительные электрические заряды, будучи положительно заряженными частицами.
Выводы
Говоря образно, «мембрана превращает клетку в «электрическую батарейку» с помощью управления ионными потоками».
Мембранный потенциал покоя образуется за счёт двух процессов:
1. Работа калий-натриевого насоса мембраны.
Новая гипотеза механизма работы Na,K-АТФазы рассматривается здесь: Механизм натрий-калиевого насоса
Работа калий-натриевого насоса, в свою очередь, имеет 2 следствия:
1.1. Непосредственное электрогенное (порождающее электрические явления) действие ионного насоса-обменника. Это создание небольшой электроотрицательности внутри клетки (-10 мВ).
1.2. Создание предпосылок для возникновения большой электроотрицательности.
2. Утечка ионов калия из клетки.
Финал
Видео: Мембранный потенциал покоя (Resting membrane potential)
Электрические потенциалы биомембран
Электрический заряд
Электрическое поле. Закон Кулона
Каждый заряженный объект образует в окружающем его пространстве электрическое поле. Электрическое поле является видом материи, посредством которой заряженные объекты взаимодействуют друг с другом. Пробный заряд, внесённый в электрическое поле другого заряда «чувствует» присутствие этого поля. Он будет притягиваться к заряду, создающему электрическое поле, или отталкиваться от него.
Закон Кулона определяет электрическую силу F, действующую между двумя точечными зарядами q1 и q2:
 |
Характеристики электрического поля
1) Напряжённость электрического поля. Каждый электрический заряд производит вокруг себя электрическое поле. Если другой заряд q внести в это поле, то на него будет действовать сила F, пропорциональная q и напряжённости электрического поля E:
 |
Напряжённость электрического поля E (или просто напряжённость) в любой точке определяется как электрическая сила F, которая действует на положительной заряд q, помещённый в эту точку:
 |
Мембранный потенциал покоя
Каждая клетка превращает часть своей метаболической энергии в электростатическую энергию. Источником электрического поля клетки является плазматическая мембрана. Существует разность потенциалов между внутренней и внешней поверхностями плазматической мембраны. Эта разность потенциалов называется мембранным потенциалом.
Разность потенциалов между внутренней и внешней средами клетки может измеряться непосредственно и довольно точно. Для этого используют микроэлектрод, представляющий собой стеклянную микропипетку с диаметром кончика до 1мкм, заполненную концентрированным раствором KCl. Микроэлектрод подключают к усилителю напряжения регистрирующего устройства. Можно измерять мембранный потенциал мышечных, нервных клеток или клеток других тканей. Другой электрод (референтный) установлен на поверхности ткани.
Когда кончик микроэлектрода находится вне клетки, его потенциал по отношению к референтному электроду равен нулю. Если конец электрода погружают в клетку, прокалывая плазматическую мембрану, разность потенциалов резко становится отрицательной. На шкале измерительного устройства регистрируется разность потенциалов между внутренней и внешней средами клетки. Эта разность потенциалов называется трансмембранной, или мембранным потенциалом.
Если клетка находится в состоянии покоя, её мембранный потенциал имеет отрицательное значение и устойчивую величину. Обычно его называют мембранным потенциалом покоя. Мембранный потенциал покоя клеток различных тканей составляет от — 55 милливольт (мВ) до —100 мВ.
При определенных физиологических условиях могут происходить изменения мембранного потенциала. Изменения его в положительном направлении называется деполяризацией плазматической мембраны. Смещение мембранного потенциала в отрицательном направлении называется гиперполяризацией.
Биофизические основы мембранного потенциала покоя
Электрические явления в плазматической мембране определяются распределением ионов между внутренней и внешней сторонами мембраны. Из химического анализа известно, что концентрация ионов внутриклеточной жидкости сильно отличается от концентрации ионов во внеклеточной жидкости. Термин «внеклеточная жидкость» имеет отношение ко всем жидкостям вне клеток (межклеточное вещество, кровь, лимфа и т.п.). В таблице представлены концентрации основных ионов в мышечных клетках млекопитающих и внеклеточной жидкости (миллимоли на литр).
 |
 |
 |
Также в поддержании мембранного потенциала покоя участвует непосредственно натрий-калий насос, выкачивая три иона натрия из клетки и закачивая лишь два иона калия. В результате мембранный потенциал покоя становится более отрицательным, чем был бы, если бы создавался только пассивным перемещением ионов через мембрану.
Потенциал действия
Если через мембрану нервной или мышечной клетки проходит кратковременный электрический ток, то мембранный потенциал подвергается последовательным изменениям, которые специфичны и уникальны для возбудимых клеток. Возбудимые ткани можно стимулировать также механическими или химическими средствами, но в экспериментальной работе, как правило, используются электрические стимулы.
 Рис. 1. Потенциал действия нервной клетки.
Рис. 1. Потенциал действия нервной клетки.
На рис. 1 показан потенциал действия нервной клетки, записанный с помощью микроэлектрода. Если к клетке прикладывают кратковременный электрический стимул, мембранный потенциал уменьшается быстро до нуля. Это отклонение характеризуют как фазу деполяризации. В течение короткого времени внутренняя среда клетки становится электроположительна по отношению к наружней (фаза реверсии мембранного потенциала, или овершут). Затем мембранный потенциал возвращается к уровню мембранного потенциала покоя (этап реполяризации) (рис.2.).
 Рис. 2. Фазы потенциала действия
Рис. 2. Фазы потенциала действия
Потенциал действия играет ведущую роль в обработке информации в нервной системе. Он имеет постоянную амплитуду, которая не является вероятностной величиной. Это имеет большое значение в обработке информации нервной системой. Кодирование интенсивности раздражения осуществляется числом потенциалов действия и частотой, с которой потенциалы действия следуют друг за другом.
Биофизические основы потенциала действия
Потенциал действия возникает из-за специфических изменений ионной проницаемости в плазматической мембране. Английский физиолог Ходжкин показал, что основной механизм потенциала действия состоит в кратковременном и очень специфическом изменении проницаемости мембраны для ионов натрия. Ионы натрия при этом поступают в клетку до момента, пока мембранный потенциал не достигнет потенциала электрохимического равновесия ионов натрия.
 Рис. 3. Изменение проницаемости мембраны для ионов натрия и калия во время потенциала действия
Рис. 3. Изменение проницаемости мембраны для ионов натрия и калия во время потенциала действия
Проницаемость мембраны для натрия при действии на клетку электрического стимула возрастает приблизительно в 500 раз и становится значительно больше, чем проницаемость мембраны для ионов калия. В клетке резко повышается концентрация ионов натрия. В результате мембранный потенциал принимает положительное значение, и поток ионов натрия в клетку замедляется.
Во время возникновения потенциала действия происходит деполяризация плазматической мембраны. Быстрая деполяризация мембраны под действием электрического стимула вызывает увеличение её проницаемости для ионов натрия. Возросшее поступление ионов натрия в клетку усиливает деполяризацию мембраны, что, в свою очередь, вызывает дальнейшее увеличение проницаемости мембраны для натрия и т.д.
 Рис. 4. Изменение состояния натриевых и калиевых каналов мембраны в зависимости от величины мембранного потенциала
Рис. 4. Изменение состояния натриевых и калиевых каналов мембраны в зависимости от величины мембранного потенциала
Распространение потенциала действия
Циклический локальный ток снижает заряд мембраны в неактивной зоне и деполяризует её. Если деполяризация достигает порогового уровня, то возрастает проницаемость мембраны для ионов натрия и возникает потенциал действия. Таким образом потенциал действия распространяется вдоль нервных и мышечных волокон с постоянной скоростью.
 Рис. 5. Распространение потенциала действия вдоль мембраны нервного волокна
Рис. 5. Распространение потенциала действия вдоль мембраны нервного волокна
Скорость распространения потенциала действия в нервных волокнах зависит от их диаметра. Она максимальна в наиболее толстых волокнах, достигая около 100 метров в секунду.
Мембранный потенциал

Потенциал покоя
Асимметричный поток катионов не будет продолжаться бесконечно: накопленный электрический потенциал (с избытком положительного заряда с левой стороны мембраны) будет противодействовать диффузии ионов калия в левый отсек. Через некоторое время поток ионов K + из правого отсека в левый сравняется по скорости с потоком из левого отсека в правый, и система достигнет равновесия. Для математического описания подобного равновесия применяют уравнение Нернста (рис. 1).
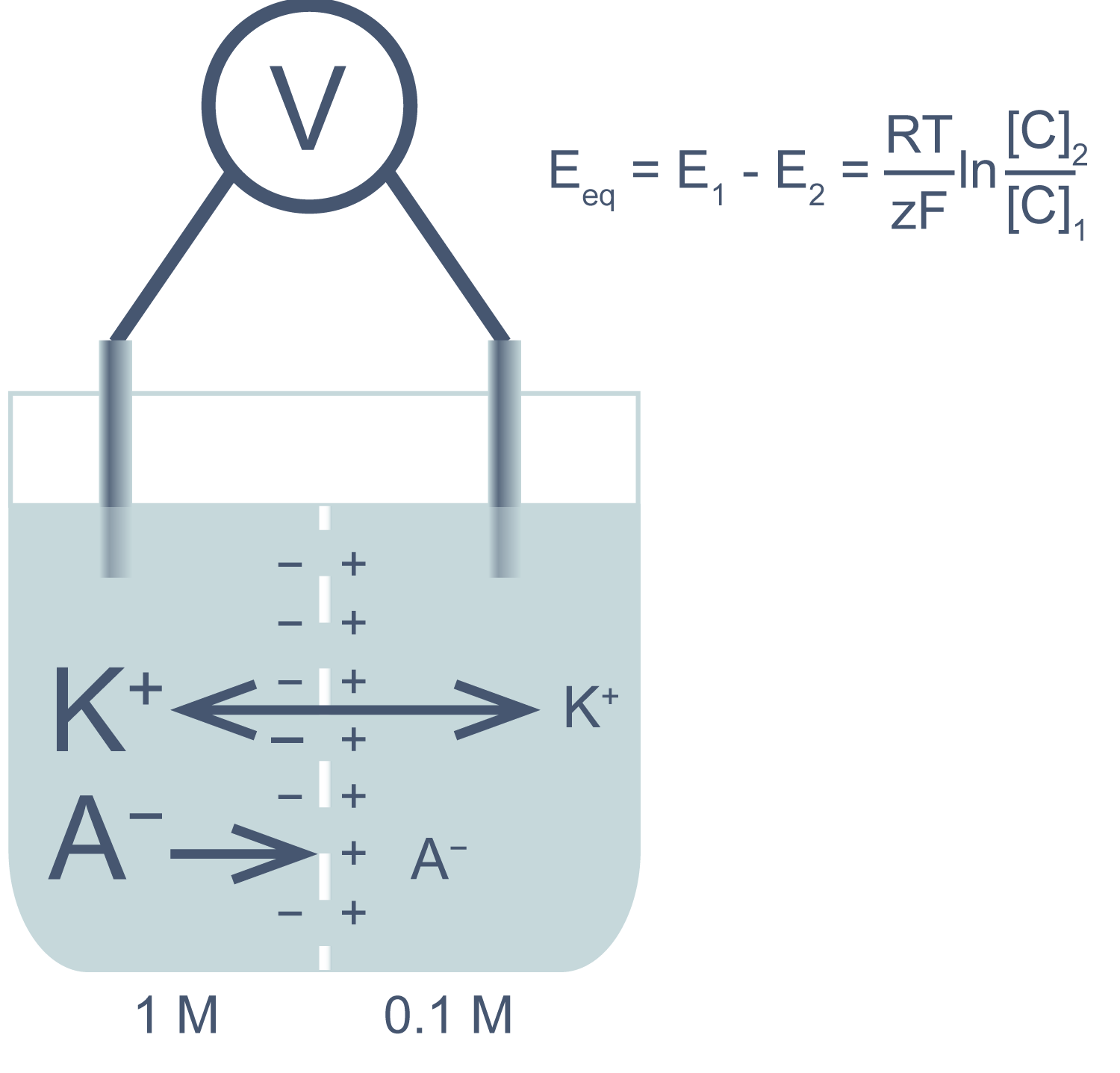
Рисунок 1 | Электрохимическая ячейка. V — вольтметр. Справа приведено уравнение Нернста, где Eeq — равновесный потенциал; E1 – E2 — разность потенциалов по обе стороны мембраны; R = 8,314 Дж/(моль·K) — универсальная газовая постоянная; T — абсолютная температура (в кельвинах); F = 96485,55 Кл·моль –1 — константа Фарадея; z — степень окисления иона (его заряд); [C]1, 2 — равновесные концентрации ионов по обе стороны мембраны.
Если принять, что равновесные концентрации ионов K + в нашем примере равны начальным, разность потенциала на мембране при 25 °C приблизительно равна –58 мВ.
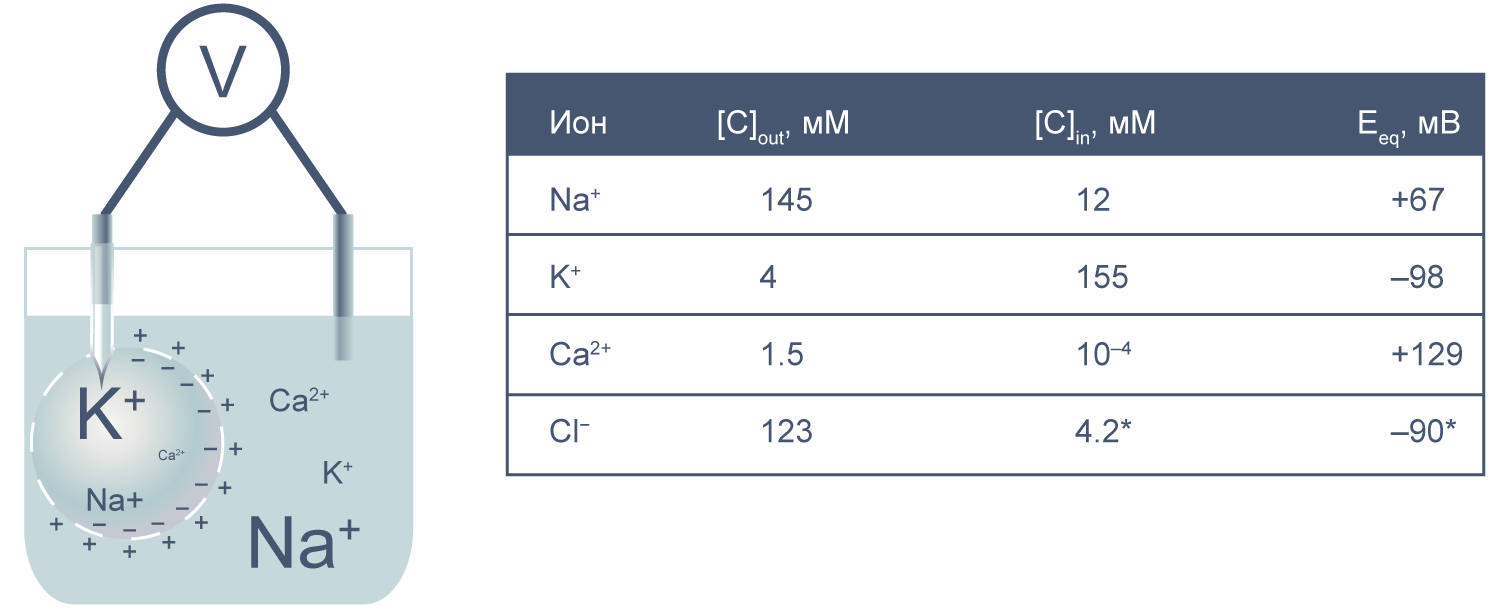
Теперь представим, что левая часть нашей электрохимической ячейки — это живая клетка, а правая — внешняя среда. Добавим к этой картине концентрации других физиологически значимых ионов. На мембране клетки также будет накапливаться электрохимический потенциал. Величину электрической составляющей мембранного потенциала измеряют относительно потенциала вне клетки, принимая его за ноль.
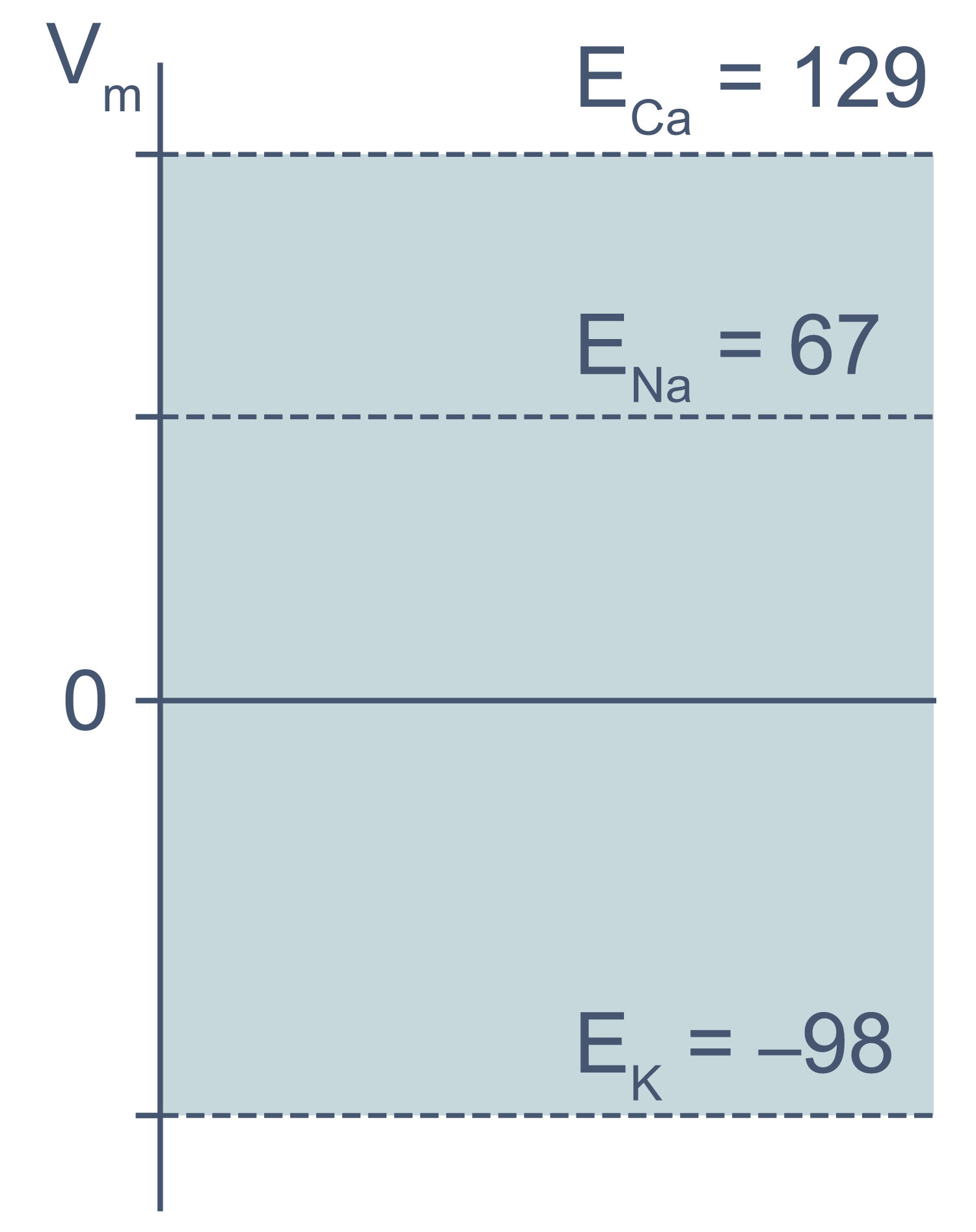
Рисунок 3 | Диапазон возможных значений мембранного потенциала от ЕK до ECa (показан голубым цветом).
Величина потенциала покоя зависит от типа клеток и равна около –30 мВ в невозбудимых клетках и около –80 мВ в возбудимых клетках (нейроны, мышечные и эндокринные клетки). Когда мембранный потенциал более отрицателен, чем потенциал покоя, говорят, что мембрана гиперполяризована, а когда он приближается к нулю или даже принимает положительные значения, говорят о деполяризации мембраны.
В общем случае мембранный потенциал можно вычислить согласно уравнению Гольдмана-Ходжкина-Катца, которое принимает в расчет все основные катионы и анионы:

где E — мембранный потенциал; R = 8,314 Дж/(моль·K) — универсальная газовая постоянная; T — абсолютная температура; F = 96485,55 Кл·моль –1 — константа Фарадея; PX — проницаемость мембраны для иона X; [C]in, out — равновесные концентрации ионов внутри и вне клетки. N.B.: для анионов внеклеточная концентрация стоит в знаменателе, а внутриклеточная — в числителе.
Что же обеспечивает проницаемость мембраны для ионов? Заряженные частицы не могут самостоятельно пересекать гидрофобный внутренний слой плазматической мембраны, и поэтому требуются специальные белки, образующие гидрофильную пору, через которую ионы могут двигаться через мембрану. Такие белки называются ионными каналами. Основной вклад в поддержание потенциала покоя вносят калиевые каналы семейств Kir (inward rectifying K + channels — калиевые каналы внутреннего выпрямления) и K2P (two—pore domain K + channels — калиевые каналы с двумя поровыми доменами, которые часто называют каналами утечки), а каналы других семейств могут обеспечивать быстрое изменение мембранного потенциала в возбудимых клетках. Каналы могут селективно пропускать определенный тип ионов, например, K + (как каналы семейства Kir), или более широкий спектр веществ, как, например, коннексины — белки щелевых контактов.
Активный и пассивный транспорт веществ через мембрану
Процессы транспорта веществ через мембрану можно классифицировать по источнику энергии для транспорта. Пассивный транспорт — это движение вещества через канал или транспортер по градиенту концентрации*, то есть за счет энергии электрохимического градиента. Таким способом через калиевые каналы пассивно движутся ионы калия, или осуществляется перенос глюкозы через транспортер GLUT4 (такой тип транспорта еще называют облегченной диффузией, а транспортеры, переносящие только один субстрат — унипортерами). Кроме пассивного транспорта, существует активный транспорт, при котором субстраты переносятся против градиента концентрации с затратой энергии, запасенной клеткой в виде АТФ (например, Na/K-АТФаза).
Некоторые транспортеры сопрягают перенос ионов или молекул против градиента концентрации с движением ионов по градиенту концентрации. Симпортеры переносят различные частицы в одном направлении (например, KCC2 — K—Cl cotransporter 2 — K/Cl котранспортер 2), а антипортеры, или обменники, — в противоположных (например, NHE-1 — Na/H exchanger 1 — Na/H обменник, участвующий в поддержании клеточного pH). Такой транспорт называется вторично-активным.
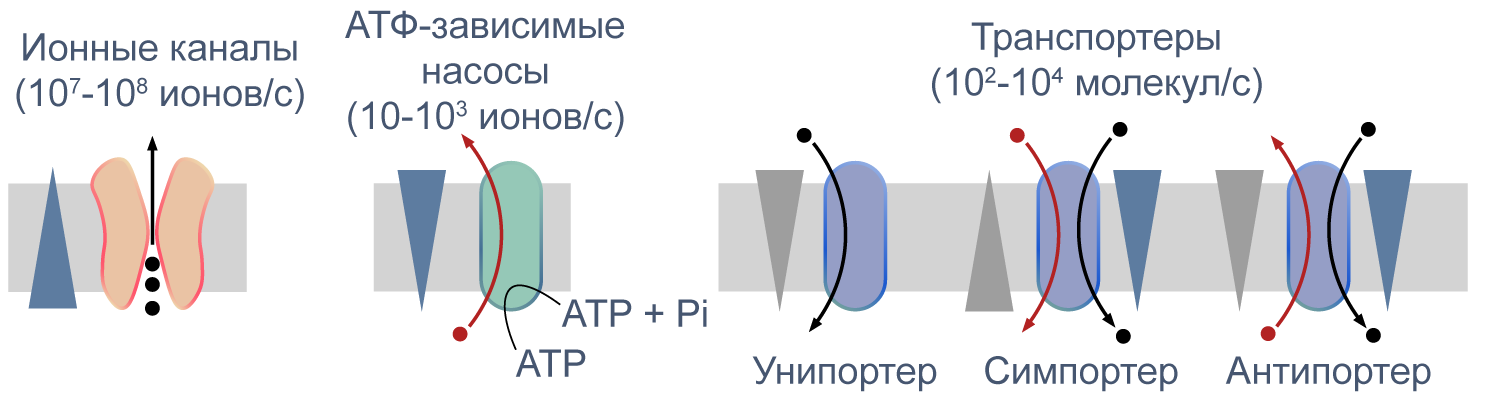
Рисунок 5 | Пассивный и активный транспорт веществ через мембрану. Треугольниками показаны концентрационные градиенты. Черными стрелками показано движение по градиенту концентрации, красными — против градиента концентрации. По [13], с изменениями.
В старой литературе можно встретить концепцию «белков-переносчиков»: до открытия молекулярной идентичности многих каналов и транспортеров (т. е. какая молекула опосредует данный ионный ток и какой ген ее кодирует) существовало представление о переносчиках как о челноках, связывающих субстраты с одной стороны мембраны, диффундирующих через мембрану и высвобождающих субстраты с другой стороны. Однако, когда стали известны аминокислотные последовательности транспортных белков, стало понятно, что все они часто содержат многочисленные трансмембранные домены и образуют сквозной путь через мембрану.
Различия между каналами, транспортерами и насосами заключаются в механизме их работы и регуляции. Каналы представляют собой более или менее селективную пору, через которую ионы могут свободно диффундировать, не вызывая конформационных изменений в белке канала. При токе через отдельный канал в 1 пА по нему проходят 6×10 6 одновалентных ионов в секунду. Транспортер, связываясь с субстратом, изменяет свою конформацию для переноса субстрата. При ко-транспорте повышается аффинность транспортера в новой конформации к второму субстрату, и субстраты переносятся через мембрану сопряженно. Насосы, также называемые АТФазами, (ауто)фосфорилируются АТФ, и это фосфорилирование значительно изменяет их конформацию и приводит к транслокации субстратов через мембрану. Электрическая проводимость насосов очень мала: Na/K-АТФаза переносит всего около 300 Na + и 200 K + в секунду.
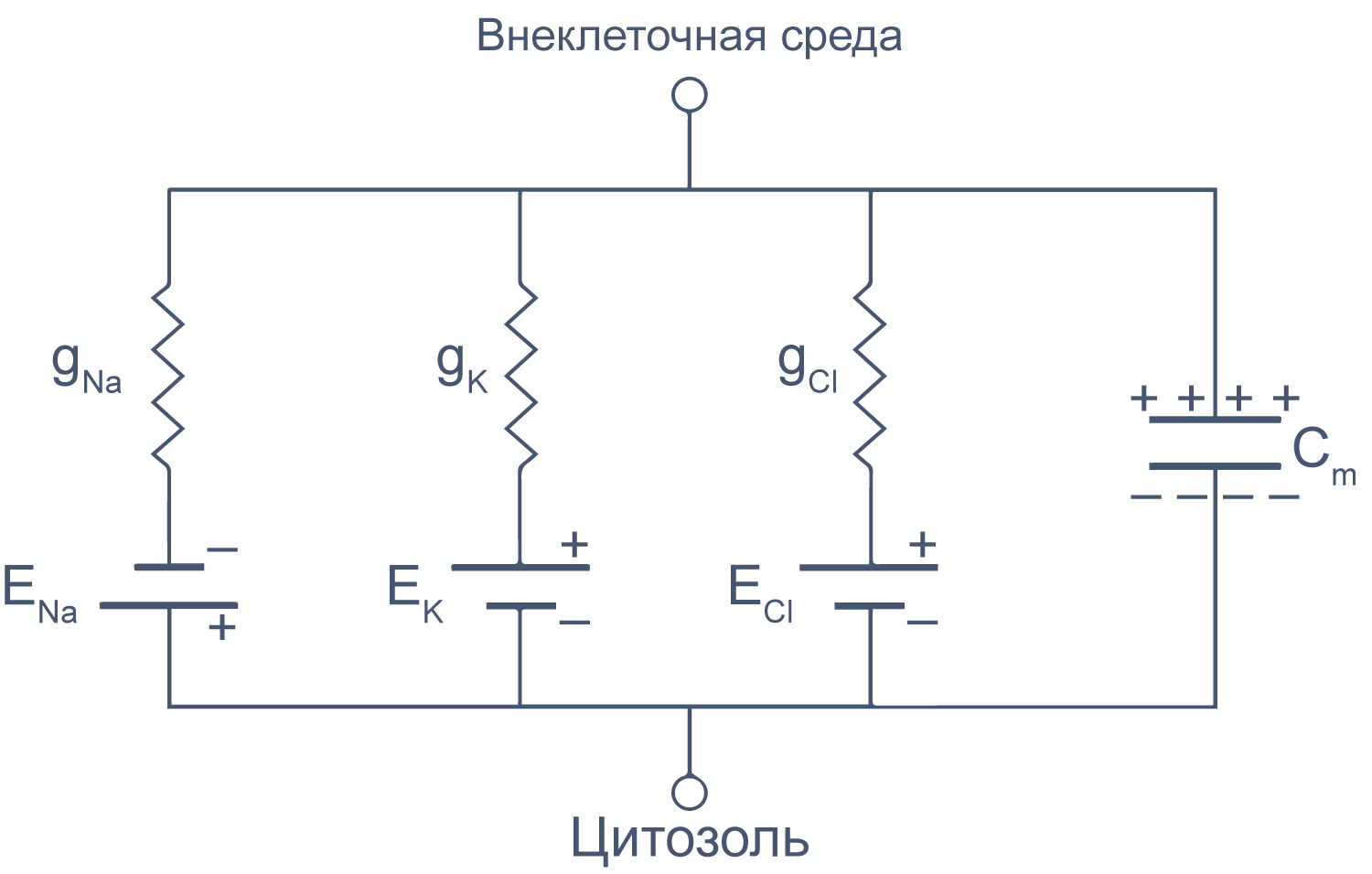
Клеточная мембрана как электрическая цепь
Липидный бислой мембраны можно представить как резистор (сопротивление) и конденсатор (емкость), соединенные параллельно. Величина сопротивления зависит от плотности каналов в мембране и их функционального состояния. Электроемкость возникает из-за разделения зарядов по обе стороны мембраны тонким слоем диэлектрика (гидрофобными хвостами фосфолипидов).
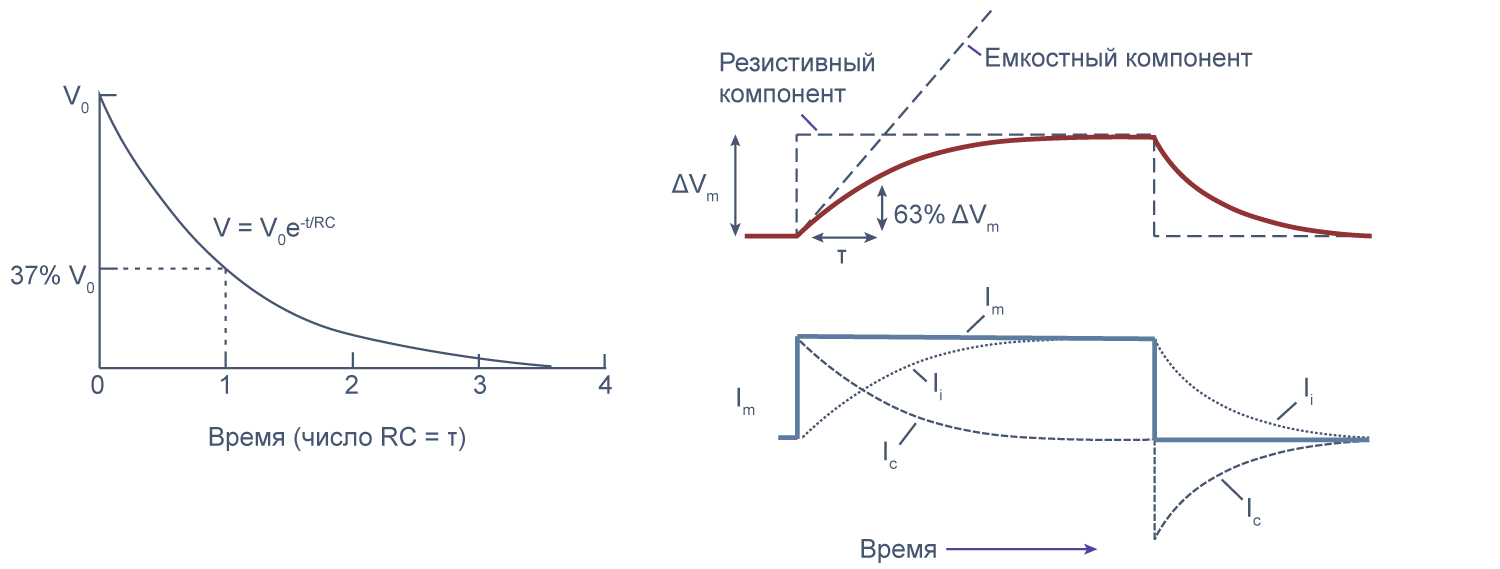
Потенциал действия
Возбудимые клетки могут быстро изменять потенциал на мембране и этим запускать внутриклеточные процессы, например, мышечное сокращение или экзоцитоз везикул. Кратковременный локальный скачок мембранного потенциала от приблизительно –80 мВ (значения потенциала покоя) до 0…+20 мВ за счет изменения проницаемости мембраны для ионов называется потенциалом действия.
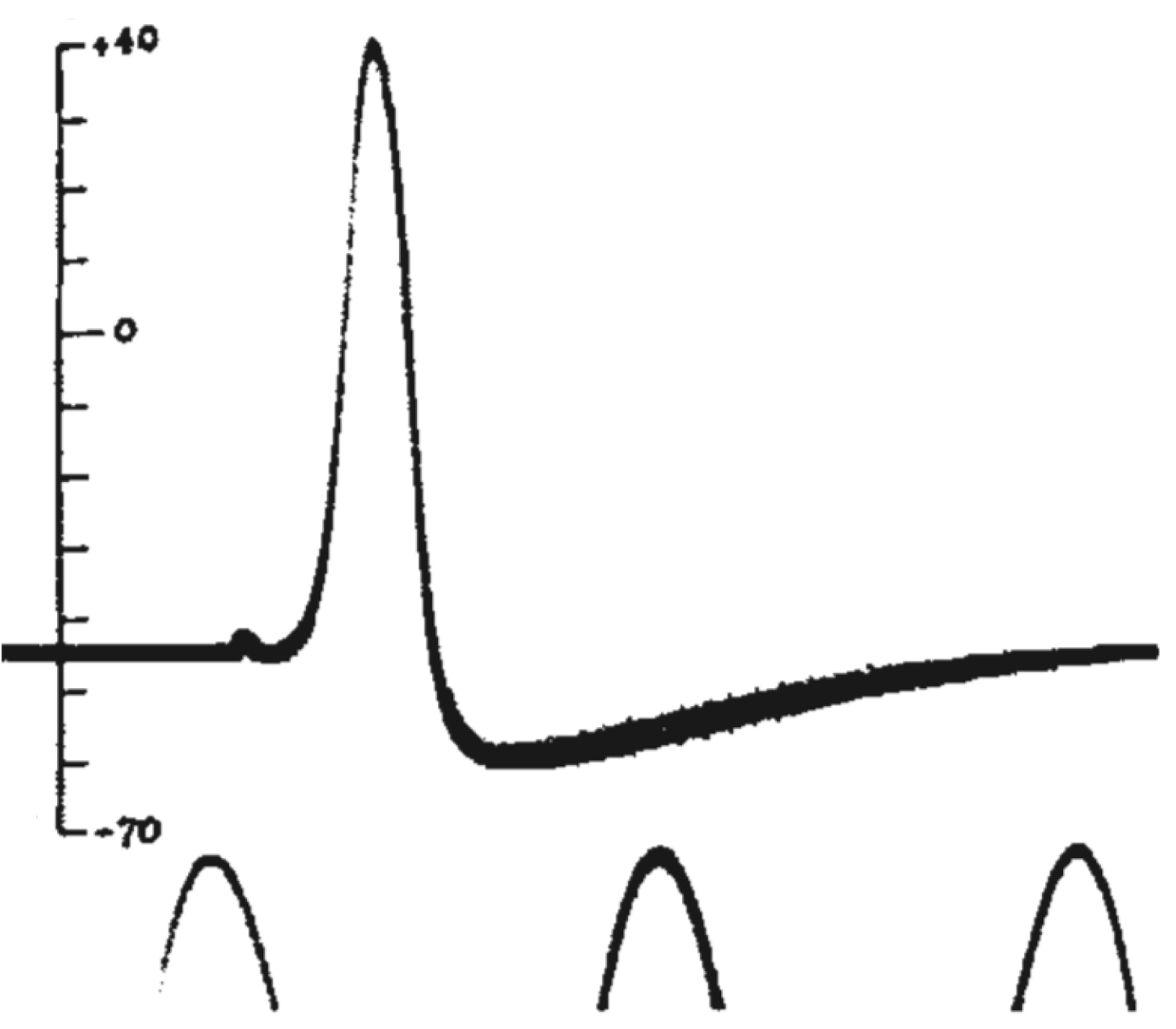
Мы видим резкую деполяризацию мембраны до положительных значений, а затем более плавное возвращение потенциала к отрицательным значениям, зачастую более отрицательным, чем величина потенциала покоя. Из этих наблюдений следовало два важных вывода: 1) потенциал действия генерируется мембраной клетки, что не было очевидно в 1930-е годы; 2) поскольку измеренная амплитуда потенциала действия была больше величины мембранного потенциала, генерация потенциала действия — это активный процесс, который нельзя объяснить временным «пробоем» в мембране (каким бы наивным такое представление нам не казалось сегодня, в 1930-е так думали многие физиологи). В том же году К. С. Коул и Х. Дж. Кертис [20] зарегистрировали кратковременное изменение проводимости мембраны с 1 мСм/см 2 до 40 мСм/см 2 при генерации потенциала действия. Эти данные также свидетельствовали о тонкой регуляции этого процесса.
После Второй мировой войны Ходжкин и Хаксли вернулись к своим исследованиям. В 1945 году они опубликовали статью с несколькими дополнительными экспериментами, подтверждающими сформулированные ранее выводы. Но и тогда ионные механизмы потенциала действия оставались неясными.
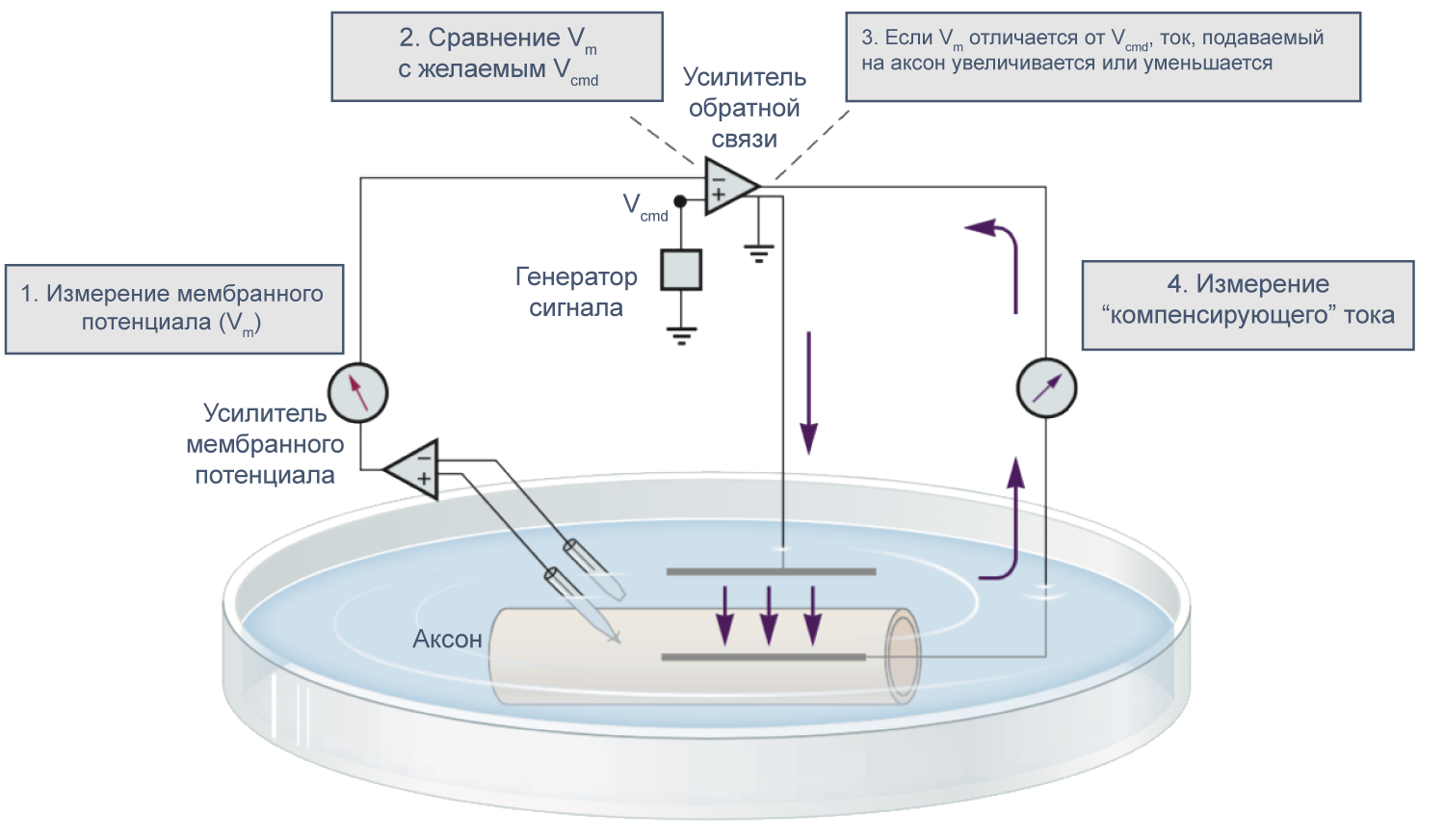
В 1970–80-х годах Эрвин Неер и Берт Сакман (Erwin Neher, Bert Sakmann) предложили метод локальной фиксации потенциала (англ. patch clamp), позволяющий работать с мелкими клетками и токами малой амплитуды и даже регистрировать активность отдельных каналов. Тем не менее, метод фиксации потенциала двумя электродами в микроэлектродной конфигурации используется и сегодня при работе с такими крупными клетками, как ооциты лягушки Xenopus laevis.
Временной ход потенциала действия
Потенциал действия в аксоне можно разделить на 1) фазу быстрой деполяризации до 2) положительных значений (овершута, от англ. overshoot), 3) фазу реполяризации, в которой потенциал возвращается к потенциалу покоя или даже до несколько более отрицательных значений — 4) следовая гиперполяризация.
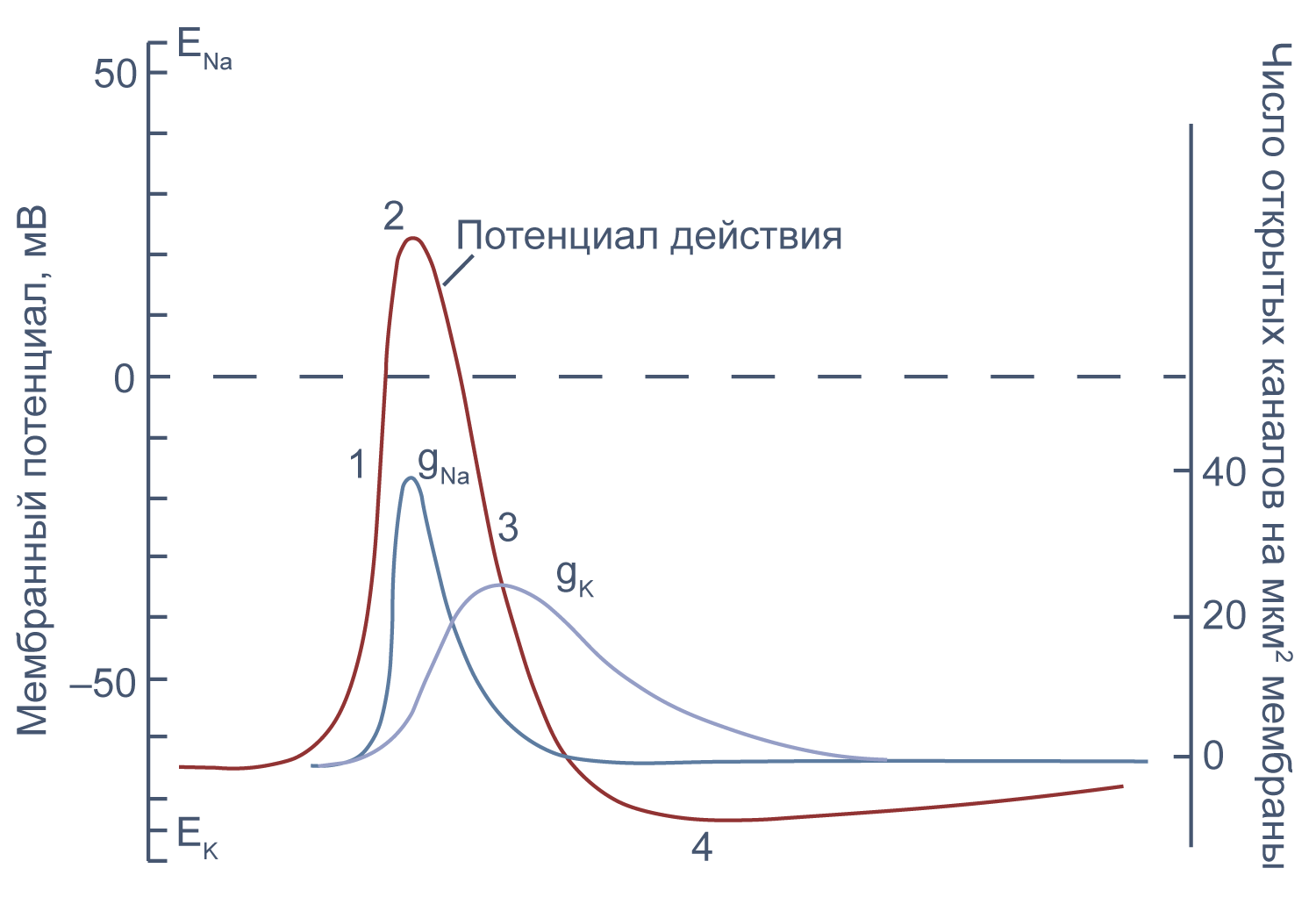
Рисунок 10 | Потенциал действия и изменения проводимости мембраны для Na + и K + в гигантском аксоне кальмара. Из [22], с изменениями.
Чтобы понять взаимосвязь ионных токов и вызванных ими изменений потенциала, рассмотрим всю цепь событий при генерации потенциала действия. Сначала мембрана деполяризуется под действием внешнего стимула: поступления в клетку катионов через лиганд-управляемые каналы, закрытия калиевых каналов или электрической стимуляции в эксперименте. Если деполяризация достигает порогового значения для потенциалзависимых натриевых каналов (Nav), они открываются, натрий по градиенту своей концентрации входит в клетку, и мембрана деполяризуется еще сильнее. Дальнейшая деполяризация влечет за собой лавинообразное открытие все новых натриевых каналов, ток через которые приводит к еще большей деполяризации. Однако эта петля положительной обратной связи не работает бесконечно: открывшись на некоторое время, натриевые каналы инактивируются и не могут открыться вновь, пока мембранный потенциал не вернется к отрицательным значениям.
Механизм инактивации был предложен еще в математической модели Ходжкина и Хаксли [23] на основании кинетических характеристик тока. Они предположили, что в натриевом канале есть три активационные частицы m и одна инактивационная частица h. Когда стала известна аминокислотная последовательность канала, выяснилось, что канал на самом деле имеет четыре гомологичных активационных домена и один инактивационный, однако один из активационных доменов срабатывает значительно медленнее остальных трех, и его влияние на кинетические характеристики тока маскируется происходящей в то же время инактивацией канала (например, [24]).
В то же время деполяризация мембраны приводит к активации потенциал-зависимых калиевых каналов (Kv), которые открываются медленнее, чем натриевые, калий выходит из клетки, и потенциал возвращается к потенциалу покоя и может даже временно стать более отрицательным: пока натриевые каналы инактивированы, потенциал становится ближе к калиевому равновесному потенциалу, и это явление называется следовой гиперполяризацией.
Генерация потенциала действия происходит по принципу «все или ничего». Если деполяризующий стимул не достиг порогового значения, потенциал действия не генерируется. Если же порог был достигнут, положительная обратная связь обеспечивает открытие всех доступных натриевых каналов, и потенциал действия достигает своей максимальной амплитуды.
Если новый стимул приходит во время или сразу после генерации потенциала действия, второй потенциал действия не возникает или обладает меньшей амплитудой, чем первый. Это явление называется рефрактерностью. Стимул, возникший в период абсолютной рефрактерности, не вызывает генерации потенциала действия, а пришедшийся на период относительной рефрактерности вызывает потенциал действия уменьшенной амплитуды, так как часть натриевых каналов все еще инактивирована.
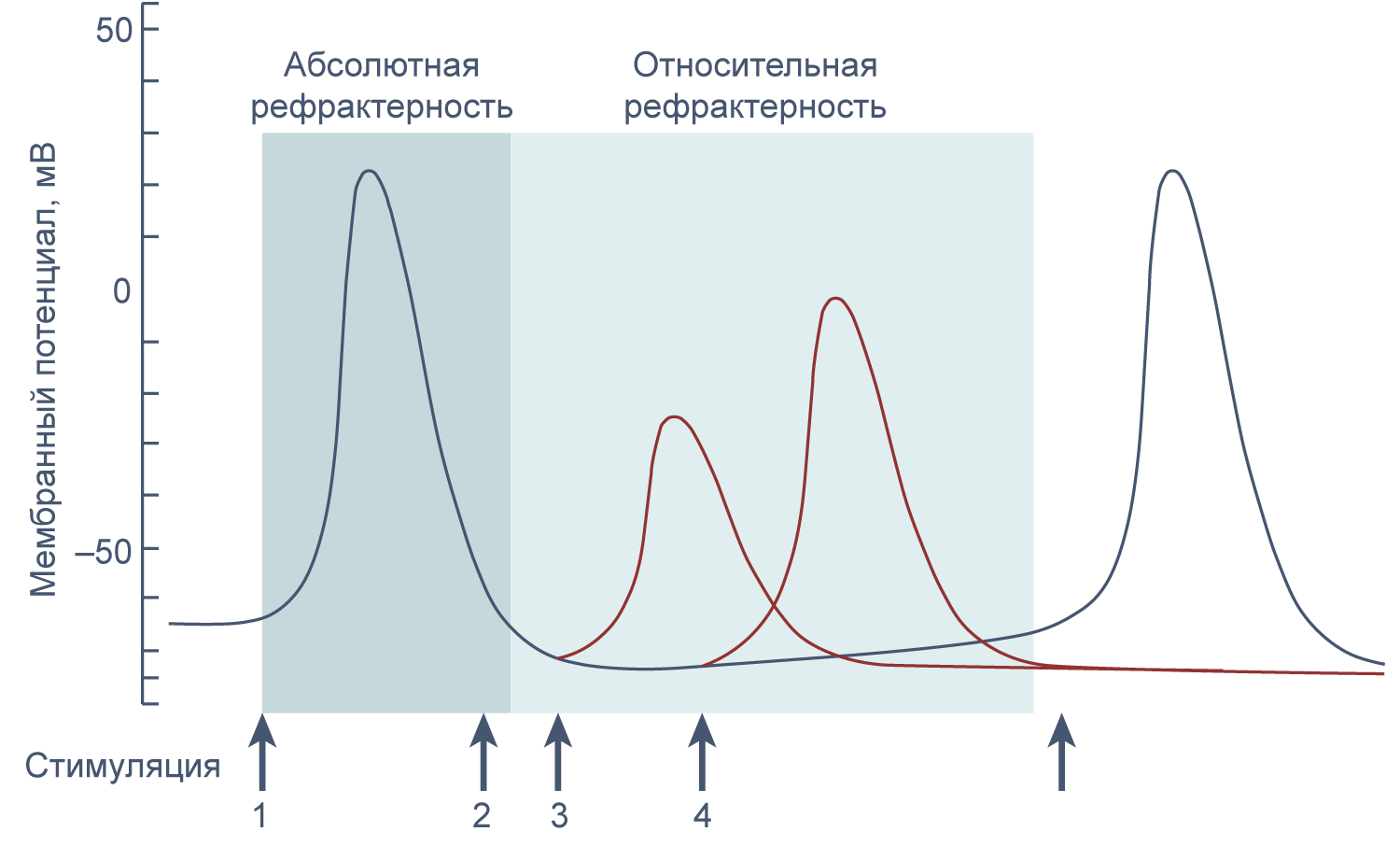
Рисунок 11 | Абсолютная и относительная рефрактерность. Стимуляция в период абсолютной рефрактерности (2) не ведет к генерации потенциала действия, а стимуляция в период относительной рефрактерности (3 и 4) приводит к генерации потенциала действия сниженной амплитуды.
Распространение возбуждения по аксону
Электрические свойства мембраны помогают понять изменения мембранного потенциала не только во времени, но и в пространстве. Пассивное, или электротоническое распространение возбуждения по мембране происходит без изменения проводимости потенциал-зависимых каналов. Для распространения потенциала действия важен как активный (изменение проводимости натриевых и калиевых каналов), так и пассивный механизмы, поскольку деполяризация, вызывающая открытие новых потенциал-зависимых натриевых каналов, должна достигнуть нового невозбужденного участка аксона, и происходит это благодаря электротоническому распространению возбуждения.
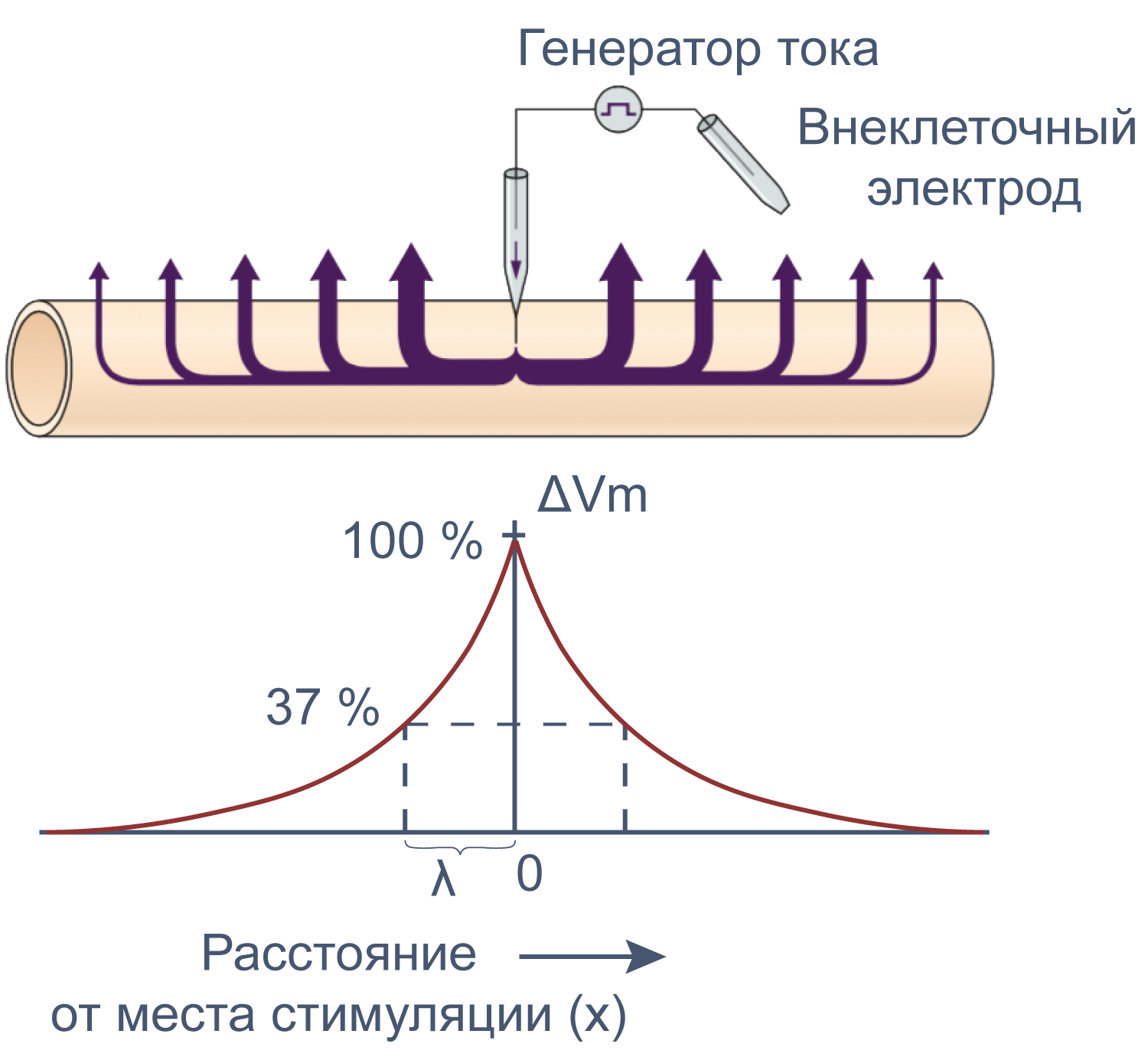
Рисунок 12 | Электротоническое распространение возбуждения. Пояснения в тексте [22].

Зависимость скорости распространения потенциала действия по аксону от его диаметра объясняет необходимость в гигантских аксонах у кальмара. Однако у позвоночных гигантских аксонов нет, и достаточная скорость проведения нервных импульсов достигается с помощью миелинизации аксонов. Миелиновые оболочки образованы специальными глиальными клетками: олигодендроцитами в центральной нервной системе и клетками Шванна в периферической. Эти клетки оборачивают аксон, образуя вокруг него плотный чехол из десятков слоев плазматической мембраны, который работает как изолятор. Под миелиновой оболочкой очень мало или почти нет натриевых каналов. Они оказываются расположены на небольших участках мембраны аксона между двумя соседними шванновскими клетками или олигодендроцитами — в перехватах Ранвье. Миелиновая оболочка увеличивает скорость проведения потенциала действия благодаря повышению сопротивления мембраны rm. При этом снижаются потери тока через мембрану, и деполяризация может пассивно распространяться на бо́льшие расстояния. Кроме того, из-за снижения емкости мембраны Cm, меньшая доля тока тратится на перезарядку мембраны. Миелинизация делает передачу потенциала действия не только быстрее, но и эффективнее, поскольку натриевые каналы располагаются только в перехватах Ранвье, благодаря чему меньшее количество ионов натрия входит в клетку и меньше энергии требуется на работу Na/K АТФ-азы для поддержания концентрационного градиента.
Потенциалы действия в других возбудимых клетках
Минималистичный натриево-калиевый потенциал действия характерен только для аксонов нейронов. В других частях нейрона и в иных типах возбудимых клеток в генерации потенциала действия принимают участие разнообразные ионные каналы, в том числе калиевые каналы других семейств и кальциевые каналы плазматической мембраны и эндоплазматического ретикулума (ЭПР является внутриклеточным депо кальция; общая концентрация Ca 2+ в нем достигает миллимолярных значений, а концентрация свободного кальция находится в микромолярном диапазоне [26] ; эти значения на несколько порядков выше, чем 100 нМ свободного Ca 2+ в цитозоле в состоянии покоя). Различия в экспрессии генов ионных каналов в различных типах возбудимых клеток порождают разнообразие потенциалов действия, различающихся ионными механизмами, длительностью (от 1,5 мс в аксонах до 500 мс в кардиомиоцитах желудочков), необходимостью внешнего стимула для генерации или наличием собственного ритма. Многие ионные каналы, не участвуя напрямую в генерации потенциала действия, влияют на возбудимость клеток и таким образом вносят вклад в разнообразие электрической активности клеток.
** Проводимость (g = 1/R) — это характеристика скорости движения любых зарядов через мембрану, а проницаемость (P) отражает, насколько легко частицы могут двигаться через мембрану независимо от того, движутся они или нет. Для ионов эти величины связаны. Например, в фазе быстрой деполяризации при потенциале действия возрастает как проводимость, так и проницаемость мембраны для ионов натрия. Однако зачастую, если проницаемость канала для какого-то иона высока из-за высокого сродства участков поры канала для этого иона, такие ионы будут двигаться через канал медленнее, и проводимость будет ниже, чем для других ионов, для которых канал менее проницаем.