перекрест двигательных путей в спинном мозге происходит
Перекрест двигательных путей в спинном мозге происходит
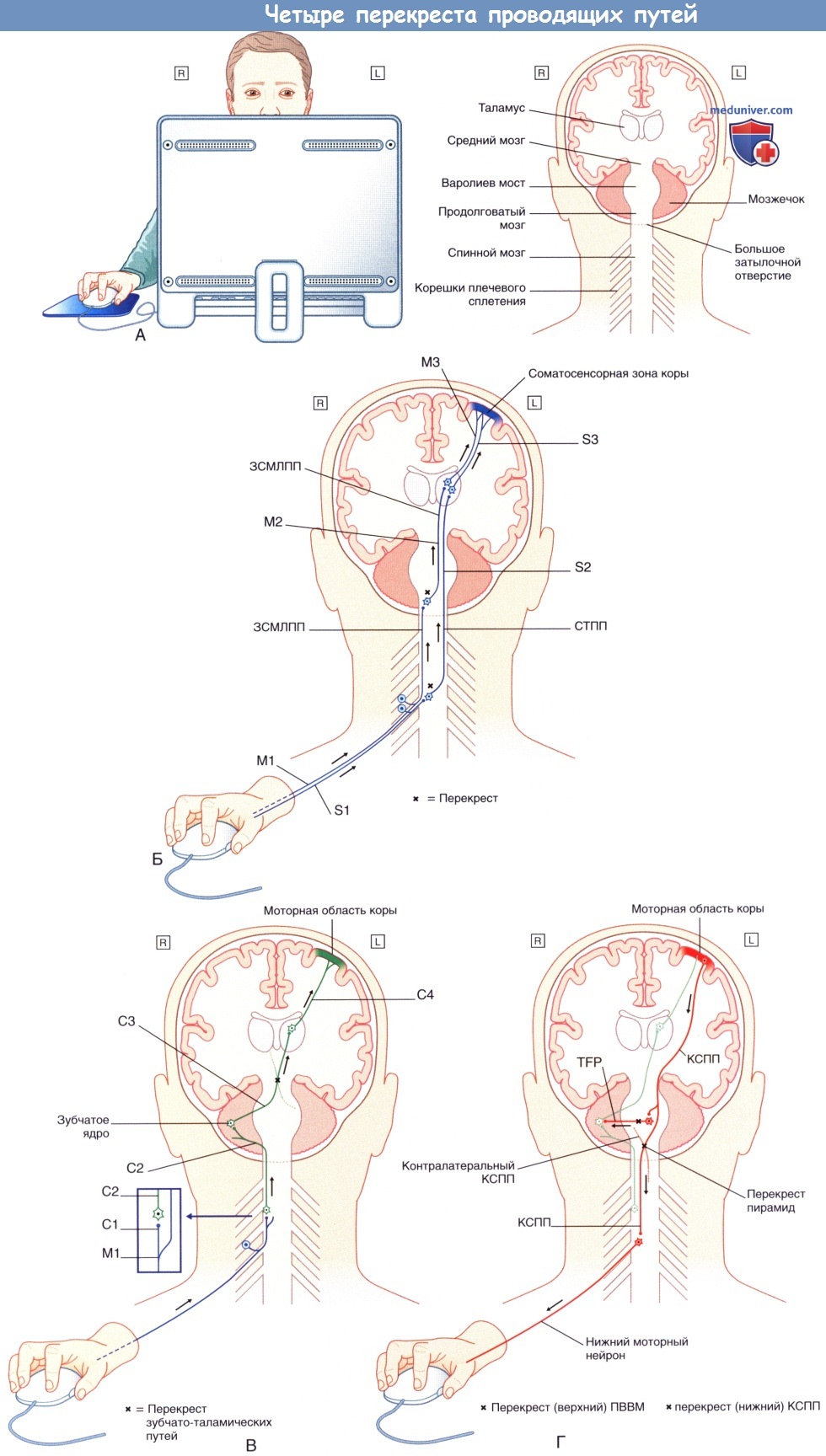 (А) Ситуационная модель: правой рукой человек кликает компьютерной мышью, в то время как его взгляд направлен в другое место. На коронарном срезе отмечены основные значимые структуры.
(А) Ситуационная модель: правой рукой человек кликает компьютерной мышью, в то время как его взгляд направлен в другое место. На коронарном срезе отмечены основные значимые структуры.
(Б) Восходящий путь сигнала. Рецепторы кожи и подлежащих тканей передают информацию в левую соматосенсорную зону коры, расположенную в постцентральной извилине, на основании которой в левой теменной доле формируется проекция взаимодействия правой руки и компьютерной мыши.
В передаче информации от кожи участвуют три последовательно расположенные группы нейронов; другие три группы нейронов осуществляют передачу информации из подлежащих тканей.
Первая группа нейронов в обоих случаях представлена нейронами первого порядка—первично-чувствующими нейронами. Такие нейроны униполярны, поскольку аксон каждого нейрона начинается из одного участка (полюса) клетки и Т-образно разделяется, чтобы обеспечить прохождение нервного импульса от тканей в центральную нервную систему.
Первично-чувствующие нейроны заканчиваются в месте соединения—синапсе—с мультиполярным (звездчатой формы) нейроном из группы нейронов второго порядка (вторичных нейронов). Аксоны нейронов второго порядка переходят на противоположную сторону, а затем направляются вверх и оканчиваются в месте контакта с мультиполярными нейронами третьего порядка (третичными нейронами), передающими сигнал в постцентральную извилину.
Первично-чувствующие нейроны, активирующиеся при воздействии на кожу кисти (S1), оканчиваются в заднем роге серого вещества спинного мозга. Чувствительные нейроны второго порядка (S2) переходят на противоположную сторону в передней белой спайке, направляются к таламусу в составе спиноталамического проводящего пути (СТПП) и переключаются на нейроны третьего порядка в зоне чувствительной коры, воспринимающей импульсы из рецепторов кисти.
Глубокую чувствительность обеспечивают нервно-мышечные веретена (мышечные веретена), расположенные в толще скелетных мышц. Первично-чувствующие нейроны мышечного веретена внутренних мышц кисти униполярны. Аксоны этих нейронов (М1) поднимаются ипсилатерально (на той же стороне) в составе задних канатиков спинного мозга, как ранее показано на рисунке ниже. Переключение нейронов первого порядка происходит в клиновидном ядре продолговатого мозга.
Аксоны мультиполярных нейронов второго порядка совершают перекрест в области чувствительного перекреста и направляются вверх (М2) через варолиев мост и средний мозг, после чего образуют синапс с нейронами третьего порядка (М3), которые направляются от таламуса к чувствительной области коры. ЗСМЛПП—заднестолбовой медиальный лемнисковый проводящий путь.
(В) Участие мозжечка. Перед подачей команды для нажатия на кнопку мыши мозг запрашивает информацию о текущем мышечном напряжении. Эта информация непрерывно поступает от мышц в полушария мозжечка на одноименной стороне. Как показано на схеме, нейроны М1 —чувствительные нейроны двойного назначения. В месте вхождения в задние канатики спинного мозга нейроны М1 формируют дополнительную ветвь С1, направляющуюся к спинно-мозжечковому нейрону, который проецируется (С2) на ипсилатеральную сторону мозжечка.
На этом уровне нейрон мозжечково-таламического проводящего пути (С3), пересекая средний мозг, направляется к контралатеральному таламусу, где следующий нейрон (С4) передает информацию к расположенной в прецентральной извилине двигательной коре, воспринимающей импульсы из рецепторов кисти.
(Г) Двигательный компонент. Мультиполярные нейроны левой части двигательной коры генерируют и передают нервный импульс по верхним мотонейронам, составляющим корково-спинномозговой проводящий путь (КСПП), который переходит на противоположную сторону в области перекреста пирамид, как ранее было показано на рисунке ниже. Нейроны корково-спинномозгового проводящего пути (КСПП) образуют синапс с нижним мотонейроном, который доносит активирующий сигнал от передних рогов серого вещества спинного мозга к мышцам-сгибателям указательного пальца и локальным стабилизирующим мышцам.
Следует отметить, что копия этой информации направляется к правому полушарию мозжечка в составе поперечных волокон варолиева моста (ПВВМ), которые образованы мультиполярными нейронами, расположенными в левой части варолиева моста.  Поперечные срезы варолиева моста.
Поперечные срезы варолиева моста.
(А) Верхняя часть варолиева моста.
(Б) Нижняя часть варолиева моста.
ВНМ, СНМ, ННМ — верхние, средние, нижние ножки мозжечка, соответственно.
КСПП — корково-спинномозговой проводящий путь.
ЗСМЛПП — заднестолбовой медиальный лемнисковый проводящий путь.  Поперечный срез продолговатого мозга.
Поперечный срез продолговатого мозга.
(А) Срез на уровне нижнего оливного ядра (НОЯ).
(Б) Срез на уровне чувствительного перекреста.
(В) Срез на уровне двигательного перекреста.
КСПП—корково-спинномозговой проводящий путь. ЗСМЛПП—заднестолбовой медиальный лемнисковый проводящий путь.
НКСПП—неперекрещенный (прямой) корково-спинномозговой проводящий путь. ННМ — нижняя ножка мозжечка.
Редактор: Искандер Милевски. Дата публикации: 9.11.2018
Вопрос 149 Двигательные проводящие пути (пирамидные и экстрапирамидные)
Эти пути можно подразделить на две группы: 1) главный двигательный, или пирамидный, путь, tractus pyramidalis (корково-ядерный и корково-спинномозговые пути), несет импульсы произвольных движений из коры головного мозга к скелетным мышцам головы, шеи, туловища, конечностей через соответствующие двигательные ядра головного и спинного мозга; 2) экстрапирамидные двигательные пути, tractus rubrospindlis, tractus ves—tibulospinalis и др., передают импульсы от подкорковых центров к двигательным ядрам черепных и спинномозговых нервов, а затем к мышцам.
К пирамидному пути, tractus pyramidalis, относится система волокон, по которым двигательные импульсы из коры большого мозга, из предцентральной извилины, от гиганто-пирамидальных нейронов направляются к двигательным ядрам черепных нервов и передним рогам спинного мозга, а от них — к скелетным мышцам. Пирамидный путь подразделяют на три части: 1) корково-ядерный — к ядрам черепных нервов; 2) латеральный корково-спинномозговой (пирамидный) — к ядрам передних рогов спинного мозга; 3) передний корково-спинномозговой (пирамидный)—также к передним рогам спинного мозга.
Корково-ядерный путь, tractus corticonuclearis, представляет собой пучок отростков гигантопирамидальных нейронов, которые из коры нижней трети предцентральной извилины спускаются к внутренней капсуле и проходят через ее колено. Далее волокна корково-ядерного пути идут в основании ножки мозга, образуя медиальную часть пирамидных путей. Корково-спинномозговые, а также корковоядерный пути занимают средние основания ножки мозга. Начиная со среднего мозга и далее, в мосту и продолговатом мозге волокна корково-ядерного пути переходят на противоположную сторону к двигательным ядрам черепных нервов: III и IV — в среднем мозге; V, VI, VII — в мосту; IX, X, XI, XII—в продолговатом мозге. В этих ядрах корково-ядерный (пирамидный) путь заканчивается, составляющие его волокна образуют синапсы с двигательными клетками этих ядер.
Латеральный и передний корково-спинномозговые (пирамидные) пути, tractus corticospinales (pyramiddles) lateralis et ventralis, также начинаются от гигантопирамидальных нейронов предцентральной извилины. Аксоны этих клеток направляются к внутренней капсуле, проходят через переднюю часть ее задней ножки, спускаются в основание ножки мозга. Далее корково-спинномозговые волокна спускаются в переднюю часть (основание) моста и выходят в продолговатый мозг, где образуют пирамиды. В нижней части продолговатого мозга часть волокон переходит на противоположную сторону и продолжается в боковой канатик спинного мозга. Эта часть пирамидных путей, участвующая в образовании перекреста пирамид (моторный перекрест), получила название латерального корково-спинномозгового (пирамидного) пути. Те волокна корково-спинномозгового пути, которые не участвуют в образовании перекреста пирамид и не переходят на противоположную сторону, продолжают свой путь вниз в составе переднего канатика спинного мозга. Эти волокна составляют передний корково-спинномозговой (пирамидный) путь. Затем эти волокна также переходят на противоположную сторону, но через белую спайку спинного мозга и заканчиваются на двигательных клетках переднего рога противоположной стороны спинного мозга.
Экстрапирамианые проворящие пути, имеют обширные связи в мозговом стволе и с корой большого мозга, взявшей на себя функции контроля и управления экстрапирамидной системой. Выделяют ряд путей экстрапирамидной системы: 1) Красноядерно-спинномозговой; 2) преддверно-спинномозговой путь; 3) корково-мостомозжечковый путь.
Красноядерно-спинномозговой путь, tractus rubrospindlis, входит в состав рефлекторной дуги, приносящим звеном которой являются спинно-мозжечковые проприоцептивные проводящие пути. Этот путь берет начало от красного ядра, переходит на противоположную сторону (перекрест Фореля) и спускается в боковом канатике спинного мозга, заканчиваясь на двигательных клетках спинного мозга.
Важным звеном в координации двигательных функций тела человека является преддверно-спинномозговой путь, tractus ves—tibulospinalis. Он связывает ядра вестибулярного аппарата с передними рогами спинного мозга и обеспечивает установочные реакции тела при нарушении равновесия. В образовании пред-дверно-спинномозгового пути принимают участие аксоны клеток латерального вестибулярного ядра, а также нижнего вестибулярного ядра преддверно-улиткового нерва. Эти волокна спускаются в латеральной части переднего канатика спинного мозга (на границе с боковым) и заканчиваются на двигательных клетках передних рогов спинного мозга.
Управление функциями мозжечка, участвующего в координации движений головы, туловища и конечностей и связанного в свою очередь с красными ядрами и вестибулярным аппаратом, осуществляется из коры большого мозга через мост по корково-мостомозжечковому пути, tractus corticopontocerebellaris. Этот проводящий путь состоит из двух нейронов. Тела клеток первого нейрона лежат в коре лобной, височной, теменной и затылочной долей. Клетки ядер моста с их отростками составляют второй нейрон корково-мостомозжечкового пути.
Перекрест двигательных путей в спинном мозге происходит
№ 194 Строение простой и сложной рефлекторных дуг. Классификация проводящих путей головного и спинного мозга.
Рефлекторная дуга — это цепь нервных клеток, обязательно включающая первый — чувствительный и последний — двигательный (или секреторный) нейроны. Наиболее простыми рефлекторными дугами являются двух- и трехнейронные, замыкающиеся на уровне одного сегмента спинного мозга. В трехнейронной рефлекторной дуге первый нейрон представлен чувствительной клеткой, который движется вначале по периферическому отростку, а затем по центральному, направляясь к одному из ядер заднего рога спинного мозга. Здесь импульс передается следующему нейрону, отросток которого направляется из заднего рога в передний, к клеткам ядер (двигательных) переднего рога. Этот нейрон выполняет проводниковую (кондукторную) функцию. Он передает импульс от чувствительного (афферентного) нейрона к двигательному (эфферентному). Тело третьего нейрона (эфферентного, эффекторного, двигательного) лежит в переднем роге спинного мозга, а его аксон — в составе переднего корешка, а затем спинномозгового нерва простирается до рабочего органа (мышца).
С развитием спинного и головного мозга усложнились и связи в нервной системе. Образовались многонейронные сложные рефлекторные дуги, в построении и функциях которых участвуют нервные клетки, расположенные в вышележащих сегментах спинного мозга, в ядрах мозгового ствола, полушарий и даже в коре большого мозга. Отростки нервных клеток, проводящих нервные импульсы из спинного мозга к ядрам и коре головного мозга и в обратном направлении, образуют пучки, fasciculi .
Пучки нервных волокон получили название проводящих путей.
В спинном и головном мозге по строению и функции выделяют три группы проводящих путей: ассоциативные, комиссу-ральные и проекционные.
Комиссуральные (спаечные) нервные волокна, neurofibrae commissurales , соединяют серое вещество правого и левого полушарий, аналогичные центры правой и левой половин мозга с целью координации их функций. Комиссуральные волокна проходят из одного полушария в другое, образуя спайки (мозолистое тело, спайка свода, передняя спайка).
Проекционные нервные волокна, neurofibrae projectiones ,
соединяют нижележащие отделы мозга (спинной) с головным мозгом, а также ядра мозгового ствола с базальными ядрами (полосатым телом) и корой и, наоборот, кору головного мозга, базальные ядра с ядрами мозгового ствола и со спинным мозгом. При помощи проекционных нервных волокон, В группе проекционных путей выделяют восходящие и нисходящие системы волокон.
Восходящие проекционные пути (афферентные, чувствительные) несут в головной мозг, к его подкорковым и высшим центрам (к коре), импульсы, возникшие в результате воздействия на организм факторов внешней среды. По характеру проводимых импульсов восходящие проекционные пути подразделяются на три группы.
1. Экстероцептивные пути несут импульсы (болевые, температурные, осязания и давления), возникшие в результате воздействия внешней среды на кожные покровы, а также импульсы от высших органов чувств (органов зрения, слуха, вкуса, обоняния).
2. Проприоцептивные пути проводят импульсы от органов движения (мышцы, сухожилия, суставные капсулы, связки), несут информацию о положении частей тела, о размахе движений.
3. Интероцептивные пути проводят импульсы от внутренних органов, сосудов, где хемо-, баро- и механорецепторы воспринимают состояние внутренней среды организма, интенсивность обмена веществ, химизм крови и лимфы, давление в сосудах.
№ 195 Проводящие пути экстероцептивных видов чувствительности. Положение проводящих путей болевой и температурной чувствительности в различных отделах спинного и головного мозга.
Экстероцептивные проводящие пути.
Проводящий путь болевой и температурной чувствительности — латеральный спинно-таламический путь, tractus spi — nothalamicus lateralis , состоит из трех нейронов. Чувствительным проводящим путям принято давать название с учетом топографии — места начала и конца второго нейрона. Например, у спинно-таламического пути второй нейрон простирается от спинного мозга, где в заднем роге лежит тело клетки, до таламуса, где аксон этого нейрона образует синапс с клеткой третьего нейрона. Рецепторы первого (чувствительного) нейрона, воспринимающие чувство боли, температуру, располагаются в коже, слизистых оболочках, а нейрит третьего нейрона заканчивается в коре постцентральной извилины, где находится корковый конец анализатора общей чувствительности. Тело первой чувствительной клетки лежит в спинномозговом узле, а центральный отросток в составе заднего корешка направляется в задний рог спинного мозга и заканчивается синапсом на клетках второго нейрона. Аксон второго нейрона, тело которого лежит в заднем роге, направляется на противоположную сторону спинного мозга через его переднюю серую спайку и входит в боковой канатик, где включается в состав латерального спинно-таламического пути. Из спинного мозга пучок поднимается в продолговатый мозг и располагается позади ядра оливы, а в покрышке моста и среднего мозга лежит у наружного края медиальной петли. Заканчивается второй нейрон латерального спинно-таламического пути синапсом на клетках дорсального латерального ядра таламуса. Здесь лежат тела третьего нейрона, отростки клеток которого проходят через заднюю ножку внутренней капсулы и в составе веерообразно расходящихся пучков волокон, образующих лучистый венец, corona radidta , достигают коры полушария большого мозга, его постцентральной извилины. Здесь они заканчиваются синапсами с клетками четвертого слоя (внутренняя зернистая пластинка). Волокна третьего нейрона чувствительного (восходящего) проводящего пути, соединяющего таламус с корой, образуют таламокорковые пучки — таламотеменные волокна, librae thalamoparietdles . Поскольку латеральный спинно-таламический путь является полностью перекрещенным проводящим путем (все волокна второго нейрона переходят на противоположную сторону), при повреждении одной половины спинного мозга полностью исчезает болевая и температурная чувствительность на противоположной стороне тела ниже места повреждения.
Проводящий путь осязания и давления , tractus spinothalamicus ventralis , передний спинно-таламический путь, несет импульсы от кожи, где лежат рецепторы, воспринимающие чувство давления и осязания, к коре головного мозга, в постцентральную извилину— место расположения коркового конца анализатора общей чувствительности. Тела клеток первого нейрона лежат в спинномозговом узле, а центральные их отростки в составе заднего корешка спинномозговых нервов направляются в задний рог спинного мозга, где заканчиваются синапсами на клетках второго нейрона. Аксоны второго нейрона переходят на противоположную сторону спинного мозга (через переднюю серую спайку), входят в передний канатик и в его составе направляются вверх, к головному мозгу. На своем пути в продолговатом мозге аксоны этого пути присоединяются с латеральной стороны к волокнам медиальной петли и заканчиваются в таламусе, в его дорсальном латеральном ядре, синапсами на клетках третьего нейрона. Волокна третьего нейрона проходят через внутреннюю капсулу (заднюю ножку) и в составе лучистого венца достигают четвертого слоя коры постцентральной извилины.
Часть волокон проводящего пути осязания и давления идет в составе заднего канатика спинного мозга вместе с аксонами проводящего пути проприоцептивной чувствительности коркового направления. В связи с этим при поражении одной половины спинного мозга кожное чувство осязания и давления на противоположной стороне не исчезает полностью, как в случае с болевой чувствительностью, а только снижается. Необходимо отметить, что не все волокна, несущие импульсы осязания и давления, переходят на противоположную сторону в спинном мозге. Этот переход на противоположную сторону частично осуществляется в продолговатом мозге.
№ 196 Проводящие пути тактильной чувствительности, их положение в различных отделах спинного и головного мозга.
Проводящий путь осязания и давления , tractus spinothalamicus ventralis , передний спинно-таламический путь, несет импульсы от кожи, где лежат рецепторы, воспринимающие чувство давления и осязания, к коре головного мозга, в постцентральную извилину— место расположения коркового конца анализатора общей чувствительности. Тела клеток первого нейрона лежат в спинномозговом узле, а центральные их отростки в составе заднего корешка спинномозговых нервов направляются в задний рог спинного мозга, где заканчиваются синапсами на клетках второго нейрона. Аксоны второго нейрона переходят на противоположную сторону спинного мозга (через переднюю серую спайку), входят в передний канатик и в его составе направляются вверх, к головному мозгу. На своем пути в продолговатом мозге аксоны этого пути присоединяются с латеральной стороны к волокнам медиальной петли и заканчиваются в таламусе, в его дорсальном латеральном ядре, синапсами на клетках третьего нейрона. Волокна третьего нейрона проходят через внутреннюю капсулу (заднюю ножку) и в составе лучистого венца достигают четвертого слоя коры постцентральной извилины.
Часть волокон проводящего пути осязания и давления идет в составе заднего канатика спинного мозга вместе с аксонами проводящего пути проприоцептивной чувствительности коркового направления. В связи с этим при поражении одной половины спинного мозга кожное чувство осязания и давления на противоположной стороне не исчезает полностью, как в случае с болевой чувствительностью, а только снижается. Необходимо отметить, что не все волокна, несущие импульсы осязания и давления, переходят на противоположную сторону в спинном мозге. Этот переход на противоположную сторону частично осуществляется в продолговатом мозге.
№ 197 Проводящие пути проприоцептивной чувствительности мозжечкового направления, их положение в различных отделах спинного и головного мозга.
По этим проводящим путям мозжечок получает информацию от расположенных ниже чувствительных центров (спинного мозга) о состоянии опорно-двигательного аппарата, участвует в рефлекторной координации движений, обеспечивающих равновесие тела без участия высших отделов головного мозга (коры полушарий большого мозга).
№ 198 Проводящие пути проприоцептивной чувствительности коркового направления, их положение в различных отделах спинного и головного мозга.
Проприоцептивный путь коркового направления также перекрещенный. Аксоны второго нейрона переходят на противоположную сторону не в спинном мозге, а в продолговатом. При повреждении спинного мозга на стороне возникновения проприо-цептивных импульсов (при травме мозгового ствола — на противоположной стороне) теряется представление о состоянии опорно-двигательного аппарата, положении частей тела в пространстве, нарушается координация движений.
№ 199 Медиальная петля, состав волокон, положение в различных отделах головного мозга.
Аксоны клеток второго нейрона проводящего пути проприоцептивной чувствительности коркового направления выходят из этих ядер, дугообразно изгибаются вперед и медиально на уровне нижнего угла ромбовидной ямки и в межоливном слое переходят на противоположную сторону, образуя перекрест медиальных петель, decussatio lemniscorum medidlium . Пучок волокон, обращенных в медиальном направлении и переходящих на другую сторону, получил название внутренних дугообразных волокон, flbrae arcuatae internae , которые являются начальным отделом медиальной петли, lemniscus medialis . Волокна медиальной петли в мосту располагаются в задней его части (покрышке), почти на границе с передней частью (между пучками волокон трапециевидного тела). В покрышке среднего мозга пучок волокон медиальной петли занимает место дор-солатеральнее красного ядра, а заканчивается в дорсальном латеральном ядре таламуса синапсами на клетках третьего нейрона.
№ 200 Двигательные проводящие пирамидные пути, их положение в различных отделах спинного и головного мозга.
Эти пути можно подразделить на две группы: 1) главный двигательный, или пирамидный, путь, tractus pyramidalis (корково-ядерный и корково-спинномозговые пути), несет импульсы произвольных движений из коры головного мозга к скелетным мышцам головы, шеи, туловища, конечностей через соответствующие двигательные ядра головного и спинного мозга; 2) экстрапирамидные двигательные пути, tractus rubrospindlis , tractus ves — tibulospinalis и др., передают импульсы от подкорковых центров к двигательным ядрам черепных и спинномозговых нервов, а затем к мышцам.
Экстрапирамианые проворящие пути , имеют обширные связи в мозговом стволе и с корой большого мозга, взявшей на себя функции контроля и управления экстрапирамидной системой. Выделяют ряд путей экстрапирамидной системы: 1) Красноядерно-спинномозговой; 2) преддверно-спинномозговой путь; 3) корково-мостомозжечковый путь.
Красноядерно-спинномозговой путь , tractus rubrospindlis , входит в состав рефлекторной дуги, приносящим звеном которой являются спинно-мозжечковые проприоцептивные проводящие пути. Этот путь берет начало от красного ядра, переходит на противоположную сторону (перекрест Фореля) и спускается в боковом канатике спинного мозга, заканчиваясь на двигательных клетках спинного мозга.
Важным звеном в координации двигательных функций тела человека является преддверно-спинномозговой путь, tractus ves — tibulospinalis . Он связывает ядра вестибулярного аппарата с передними рогами спинного мозга и обеспечивает установочные реакции тела при нарушении равновесия. В образовании пред-дверно-спинномозгового пути принимают участие аксоны клеток латерального вестибулярного ядра, а также нижнего вестибулярного ядра преддверно-улиткового нерва. Эти волокна спускаются в латеральной части переднего канатика спинного мозга (на границе с боковым) и заканчиваются на двигательных клетках передних рогов спинного мозга.
Управление функциями мозжечка, участвующего в координации движений головы, туловища и конечностей и связанного в свою очередь с красными ядрами и вестибулярным аппаратом, осуществляется из коры большого мозга через мост по корково-мостомозжечковому пути, tractus corticopontocerebellaris . Этот проводящий путь состоит из двух нейронов. Тела клеток первого нейрона лежат в коре лобной, височной, теменной и затылочной долей. Клетки ядер моста с их отростками составляют второй нейрон корково-мостомозжечкового пути.