поля головного мозга таблица
Поля головного мозга таблица
Наиболее широко используют карту полей Бродмана, на которой кора разделена на 44 цитоархитектонические зоны (его схема была расширена до 52 полей, однако не все поля использовали). Большая часть полей представлена на рисунке ниже, однако четких границ между этими полями не существует. (Эти числа часто используют для обозначения функциональных областей, хотя Бродман отвергал подобную корреляцию.)
На рисунке ниже разным цветом выделены три основные первичные сенсорные зоны (соматическая, зрительная и слуховая) и одна первичная моторная зона. Рядом с каждой первичной сенсорной или моторной зоной расположена ассоциативная кора—унимодальная ассоциативная зона (той же модальности). Остальная часть новой коры представлена мультимодальными (полимодальными) ассоциативными зонами, к которым подходят волокна от многих унимодальных ассоциативных зон (например, от зон тактильной и зрительной чувствительности, зрительной и слуховой) и других мультимодальных или паралимбических областей.
 Цитоархитектонические поля Бродмана. Окрашенные зоны:
Цитоархитектонические поля Бродмана. Окрашенные зоны:
а) Моторная (красная):
4 — первичная моторная кора
6 — на медиальной поверхности, дополнительная моторная зона
6 — на латеральной поверхности, премоторная кора
б) Сенсорная (голубая):
3/1/2 — первичная соматосенсорная кора
40 — вторичная соматосенсорная кора
17 — первичная зрительная кора 18,
19 — ассоциативная зрительная кора
41, 42 — первичная слуховая кора*
22 — ассоциативная слуховая кора
(*Первичную слуховую кору не всегда можно увидеть сбоку, так как она полностью расположена на верхней поверхности верхней височной извилины.)
Изучение функциональной анатомии. Термином коннектом обозначают «полную карту нейронных связей, охватывающих все функции мозга». Однако для создания законченной функциональной карты человеческого мозга требуется объединить эмпирические данные со структурными связями, при этом многое все еще остается неизвестным. Современные подходы создают уникальные возможности для достижения этой цели с помощью новых возможностей обработки и хранения данных, нейрофизиологических исследований и магнитно-резонансной томографии (МРТ), позволяющих получить изображения головного мозга живого человека.
Новые достижения в понимании устройства мозга ознаменовались смещением приоритета с отдельных зон коры на рассмотрение всех отделов и взаимосвязей как единого целого. Были разработаны новые теоретические и методологические схемы, позволяющие описывать и прогнозировать сложную системную динамику путем использования сетевого анализа и математических методов, основанных на теории графов. В сетевых моделях используют совокупности «элементарных» корковых единиц и их взаимодействия, чтобы показать появление функционирующих участков в динамике или «поймать мозг в действии».
Эти модели остаются ограниченными известными взаимосвязями между зонами коры, а существование некоторых взаимосвязей было предположено по результатам исследований у приматов. Однако данные модели позволяют предположить наличие взаимосвязей или проводящих путей, существование которых структурно не доказано, на основании типа реакции. Несмотря на прогресс в исследовании проводящих путей и взаимосвязей в коре при лучевой диагностике мозга живого человека благодаря использованию нейрорентгенологических методов и математического моделирования, внедрение новых и продолжение использования «старых» техник изучения нейроанатомии необходимо для получения структурных свидетельств существования этих проводящих путей и возбуждающих нейронных систем.
Для «определения и локализации» функций головного мозга используют преимущественно две методики. В основе обеих лежит регистрация локального усиления кровотока в ответ на повышенную потребность мозга в кислороде в результате увеличения нейронной активности.
1. Позитронная эмиссионная томография (ПЭТ). С помощью позитронной эмиссионной томографии (ПЭТ) измеряют потребление кислорода после введения в вену предплечья воды, меченной кислородом-15 ( 15 O). 15 O — излучающий позитроны изотоп кислорода; в крови позитроны реагируют с окружающими электронами и испускают у-лучи, которые регистрируют детекторы γ-лучей. Для измерения уровня потребления глюкозы используют также 18-фтордезоксиглюкозу ( 18 F-дезок-сиглюкоза). Нейроны захватывают 18 F-дезоксиглюкозу в той же степени, что и глюкозу.
Для правильной интерпретации данных ПЭТ требуется выполнение вычитания изображений и усреднения изображений, описанных под рисунком ниже. Аналогичная методика извлечения сигнала описана для функциональной МРТ (фМРТ).
При проведении специализированных исследований (например, при определении суммарной функции рецепторов) используют меченные изотопом химические вещества: меченный радиоизотопом дофамин в полосатом теле при болезни Паркинсона, меченный радиоизотопом серотонин в стволе мозга и коре больших полушарий при депрессии, меченую ацетилхолинэстеразу при болезни Альцгеймера.
2. Функциональная магнитно-резонансная томография (фМРТ). При фМРТ не требуется введения дополнительных веществ. Метод основан на различии в магнитных свойствах оксигенированной и неоксигенированной крови. Если локального усиления кровотока более чем достаточно для покрытия потребности в кислороде, повышается коэффициент отношения оксигемоглобина к дезоксигемоглобину, что ведет к образованию MPT-сигнала. Функциональные и структурные взаимосвязи можно выявить при взаимном изменении или колебании интенсивности фМРТ-сигнала в различных корковых зонах даже при отсутствии «прямых» кортикальных связей. Приведенные ниже данные были получены с помощью функциональных методов исследования, клинических наблюдений и результатов экспериментов на животных.
 Вычитание изображений и усреднение изображений при позитронной эмиссионной томографии (ПЭТ).
Вычитание изображений и усреднение изображений при позитронной эмиссионной томографии (ПЭТ).
Верхний. Контрольное среднее изображение получено у пациента в покое. Захват 15 O происходит во всех отделах коры и подкоркового серого вещества.
Левое изображение получено у того же пациента, следящего за движущимися по экрану точками.
Высокий уровень фоновой активности скрывает результат исследования. Правое изображение получено вычитанием контрольного изображения и позволяет увидеть повышенную активность зрительной коры при выполнении зрительной задачи.
Средний. Аналогичную задачу выполняли другие четыре пациента. Вычитание фонового «шума» позволило выявить значительные различия между пациентами.
Поскольку размеры мозга у людей варьируют, активность мозга у этих пятерых пациентов была наложена на общий, «средний» мозг (хотя мозг на всех рисунках изображен одинаково). Нижний. Усредненное значение пяти исследований указывает на среднюю разницу в этой группе.
Редактор: Искандер Милевски. Дата публикации: 22.11.2018
Цитоархитектонические поля Бродмана в полушариях головного мозга
Нейроны  коры больших полушарий играют роль улавливания, приема и передачи многообразных видов раздражений. Они передают импульсы в центральную нервную систему и иные ядра анализаторов.
коры больших полушарий играют роль улавливания, приема и передачи многообразных видов раздражений. Они передают импульсы в центральную нервную систему и иные ядра анализаторов.
Иван Петрович Павлов описывал полушария головного мозга как скопление корковых анализаторов, где рождаются ответные реакции, влияющие на активность организма.
История изучения
В Германии в 1909 году врач-ученый Корбиниан Бродман впервые построил и описал карты цитоархитектонических полей Бродмана. Спустя 10 лет О. Фогт и Ц. Фогт изучили и описали больше сотни миело-архитектонических областей в полушариях мозга.
В результате в государственном научном центре неврологии И. Н. Филимоновым и С.А. Саркисовым выпущена карта полушарий мозга по Бродману, в которой описаны 47 полей.
Поля Бродмана чаще всего используются при изучении нейронной организации головного мозга и ее функционирования. Разделение той или иной области мозга к конкретному разделу полей проводилось на основе гистологического анализа — цветовой окраске по Нисслю.
Группы полей головного мозга по Бродману
Описание полей Бродмана в коре головного мозга по зонам:
Схема полей Бродмана:

Центры первой сигнальной системы
Первая сигнальная система числится как у Homo sapiens, так и у других живых существ. Она выполняет функции понимания и анализа раздражений, исходящих из окружающего мира и проявляющиеся в форме ощущений и представлений.
Центры первой сигнальной системы располагаются в ядрах анализаторов сенсорной чувствительности, двигательных анализаторах, слуховых, кожных, зрительных и обонятельных.
Обозначаются в верхней и нижней теменной области мозга, в прецентральной и надкраевой борозде, а также в толще латеральной борозды (слуховые ядра).
Назначение данной системы представлено в таблице полей Бродмана:
| Поля по Бродману | Ядра анализатора | Функции |
|---|---|---|
| 1, 2, 3, 5, 7 | Корковые | Ответственно за восприятие температуры и боли, а также осязательные ощущения. Проводящие аффективные пути, тянущиеся к коре большого мозга, пересекаются в районе спинного и продолговатого мозга. Из-за этого функции любого из полушарий контролируются диаметрально инверсной частью тела. |
| 6 и 4 | Моторные | Здесь выделяются нейроны, реакции от которых контролируют мышцы нижней части тела и лицевые. |
| 8 | Черепа и глаз | Ядра, которые контролируют движения головы и глаз. |
| 40 | Моторные | Контролируют произвольные целенаправленные движения животного или человека. |
| 18, 17 и 19 | Зрительные | Отвечают за зрительную память, восприятие образов и ориентировку в незнакомых местах. |
| 7 | Кожные | Выполняет тактильные функции распознавания объектов и поверхностей на ощупь и других видов кожной чувствительности. |
| 41, 42 и 52 | Слуховые | Восприятие и запоминание звуков извне. |
| 43 | Обонятельное | Филогенетически самая старая область коры мозга. Обеспечивает функцию восприятия и запоминания запахов. Тесно связана с вкусовыми ощущениями. |
На картинке изображены поля коры больших полушарий по Бродману и их функции:

Вторая сигнальная система
Вторая сигнальная система существует исключительно у Homo sapiens, ее возникновение объясняется речевыми способностями и мышлением.
Размещается вторая сигнальная система в ядрах моторного анализатора написанного текста, речедвигательного анализатора, понимания вербальной и невербальной речи. Ядра второй системы раскинулись в задних и центральных областях нижележащей лобной извилины, а также в вышележащей височной борозде и внизу теменной доли.
По ядрам определяется назначение второй сигнальной системы.
Ядро восприятия письменности включает в себя поле 40, оно отвечает за изображение и понимание написанных букв. Контролирует также движения рук, повороты черепа и глаз, тонкую моторику.
Речедвигательная область состоит из поля 44 — это речевой центр Брокка, а также 45, связанное с музыкальным восприятием. Это ядро взаимосвязано с областями движений, так как в речевом процессе играет роль моторика многих мышц языка и рта. Выполняет функции устного воспроизведения звуков, слов и фраз.
Слуховое ядро устной речи состоит из поля 42 и 22, контролирует задачу распознавания и понимания устных звуков речи.
Ядро письменной речи — поле 29, обеспечивает функции анализа, восприятия и понимания написанного текста и чтения.
Поля головного мозга таблица
Ожидайте
Перезвоните мне

Ваш персональный менеджер: Екатерина
Ответственная и отзывчивая! 😊

Аннотация: 3 блока мозга как структурно-функциональная модель.
Статья:
3 блока мозга как структурно-функциональная модель. Признаки нарушения в развитии блоков мозга
В начале 20 века А.Р. Лурия разделил (условно) мозг человека на 3 функциональных блока, взаимодействие которых необходимо для любой психической деятельности.

1-й блок мозга преимущественно ответствен и за эмоциональное «подкрепление» психической деятельности (переживание успеха – неуспеха).
Этот блок мозга участвует в организации внимания, памяти, эмоционального состояния (особенно страх, боль, удовольствие, гнев), перерабатывает разнообразную информацию о состоянии внутренних органов и регулирует эти состояния, а так же поддерживает общий тонус ЦНС.
Все, что происходит с мамой во время беременности (болезни, психотравмы, прием лекарств и т.д.) откладывает свой отпечаток на формирование 1 блока мозга.

2 блок – приема, переработки и хранения информации – формируется от 3х до 7 лет и включает в себя основные анализаторные системы: зрительную, слуховую и кожно-кинестетическую, корковые зоны которые расположены в задних отделах больших полушарий головного мозга.
Поражение третичных зон приводит к нарушению комплексного синтеза раздражений, поступающих от разных анализаторов, что проявляется в нарушении ориентировки в пространстве.
Поля головного мозга таблица
а) Первичная моторная кора. Первичная моторная кора (поле 4) — полоска агранулярной коры в прецентральной извилине. Она дает начало 60-80% (количество варьирует) волокон корково-спинномозгового пути (КСП). Остальная часть волокон отходит от премоторной, поясной и добавочной моторной зон, а также теменной коры, что показано в отдельной статье на сайте. Наибольшую плотность окончаний КСП в спинном мозге наблюдают в зонах, иннервирующих дистальные мышцы конечностей.
Части тела противоположной стороны соматотопически представлены в коре инвертированными, за исключением лица, а относительно крупные области занимают кисть (играет важную роль в контроле движений пальцев), околоротовая область и язык. Зону кисти обычно можно определить по направленному кзади возвышению в 6-7 см от верхнего края полушария.
Ипсилатералъные части тела также имеют соматотопическую организацию; ипсилатеральные моторные нейроны составляют лишь 10% волокон пирамидного пути, которые не переходят на противоположную сторону и не иннервируют дистальные мышцы конечностей.
При целенаправленной стимуляции моторной коры человека установлено, что клеточные колонны регулируют направление движения. Первичная моторная кора «синтезирует» двигательные команды, но не является местом их возникновения. От нее волокна в составе КСП направляются в спинной мозг и широко разветвляются по мере достижения места своего окончания. Для того чтобы взять, например, ручку в руку, требуется:
(а) умеренное сокращение мышцы, противопоставляющей большой палец, как основной движущей мышцы;
(б) сокращение с той же интенсивностью отдела глубокого сгибателя пальцев, от которого отходит сухожилие к дистальной фаланге указательного пальца;
(в) более слабое сокращение мышцы, приводящей большой палец, и
(г) короткого сгибателя большого пальца.
Удержание всей руки при любом типе манипуляций — функция премоторной коры, отражающая важность бессознательной регуляции положения тела при совершении произвольных движений. Более крупные моторные зоны коры формируются при «объединении» близлежащих нейронных колонн на основе их функций и создании сложных последовательностей движений.
1. Пластичность моторной коры. Некрупные поражения моторной коры у обезьян и низших млекопитающих приводят первоначально к параличу соответствующей части тела, а затем в течение нескольких дней (иногда часов) — к быстро прогрессирующему восстановлению функции. Восстановление может быть обусловлено изменением свойств клеточных колонн, расположенных рядом с местом поражения, которые берут на себя утраченную двигательную функцию. Увеличение двигательной зоны участка коры возможно путем локального введения антагонистов ГАМК в кору. Расширение моторной зоны на уровне спинного мозга заранее обеспечено обширными зонами перекрытия от поля 4 до колонок двигательных нейронов серого вещества переднего рога, однако степень пластичности здесь ниже, чем в коре.
Взаимосвязи между КСП (и другими нисходящими путями) и двигательными нейронами в спинном мозге возникают посредством вставочных нейронов. Во вставочных нейронах происходит объединение сенсорной и корковой информации, что сопровождается специфической и организованной активацией пулов двигательных нейронов и сокращением иннервируемых ими мышц.
2. Источники афферентных путей к первичной моторной коре:
— Моторная кора противоположной стороны через мозолистое тело. Наиболее прочные комиссуральные связи возникают между соответствующими колоннами клеток, иннервирующими мышцы живота и спины. Это вполне понятно, поскольку данные группы мышц обычно сокращаются одновременно с двух сторон, поддерживая вертикальное положение головы и туловища. Самые слабые комиссуральные связи имеются между колоннами клеток, контролирующими дистальные группы мышц конечностей, движения в которых слева и справа происходят независимо друг от друга.
— Соматосенсорная кора. Кожные колонны клеток в полях 1, 2 и 3 отдают кпереди короткие ассоциативные волокна (взаимосвязи с зоной кисти особенно многочисленные; эти волокна короткие, так как зоны, отвечающие за кисть в сенсорной и моторной коре, обычно занимают соседние стенки центральной борозды). Проприоцептивные клеточные колонны получают афферентые волокна от аннуло-спиральных окончаний мышечных веретен; они отдают короткие ассоциативные волокна к соответствующим двигательным колоннам, активируя рефлексы растяжения с длинной дугой.
— Противоположное зубчатое ядро. Мозжечок принимает участие в выборе нужных мышц-синергистов, а также времени и силы их сокращений.
— Дополнительная моторная зона (ДМЗ).
 (А) Предполагаемая функциональная организация сегмента запястье-кисть в первичной двигательной коре (М1) у обезьян и других приматов.
(А) Предполагаемая функциональная организация сегмента запястье-кисть в первичной двигательной коре (М1) у обезьян и других приматов.
Несмотря на то, что М1 имеет в целом соматотопическую организацию, локальная соматотопическая структура разбита в виде мозаики на радиальные ряды нейронов, инициирующих небольшие специфические движения.
Миниколонны, отвечающие за движения отдельных пальцев, могут примыкать к колоннам запястья, локтя или плеча, а подгруппы этих миниколонн объединены по функции образования более сложных последовательностей движений, таких как хватание, вытягивание руки или защита рукой от удара.
(Б) Пространственная организация моторных нейронов коры приматов, контролирующих движения мышечных групп.
От медиальной стороны к латеральной они располагаются в следующей последовательности: стопа, голень, плечо, кисть и голова (голубой).
В каудальном отделе М1 лежат двигательные нейроны, взаимодействующие непосредственно с двигательными нейронами (спинного мозга) и отвечающие за высокоточные движения, необходимые для выполнения тонких двигательных навыков.
Эти нейроны расположены в медиально-латеральной последовательности от проксимальных (красный) до дистальных (желтый) групп мышц. Авторы обозначают эту зону как «новую М1», подотдел «старой М1» (голубая) для того, чтобы отразить недавнее возникновение сложной двигательной активности и эволюционно «нового» отдела моторной коры.
б) Премоторная кора. Премоторная кора (поле 6 на латеральной поверхности полушария) примерно в 6 раз крупнее первичной моторной коры. Она получает когнитивные импульсы от лобной доли при намерении произвести движение, а также от теменной доли (поле 7), передающей огромное количество тактильной и пространственно-зрительной информации. Максимальная ее активация происходит, когда последовательность движений происходит в ответ на зрительные или соматосенсорные раздражители, такие как приближение к объекту, полностью находящему в поле зрения, или идентификация объекта, находящегося вне поля зрения путем ощупывания. Активация премоторной коры, как правило, двусторонняя. Одним из объяснений служит межполушарная передача двигательных программ через мозолистое тело.
Следует также принимать во внимание обширные связи премоторной коры с ядрами ствола мозга, дающими начало ретикуло-спинномозговым путям (и небольшому пути к КСП). Поражения, затрагивающие премоторную кору у человека, встречают редко, они характеризуются нестабильностью положения в противоположном плече и бедре. Таким образом, важной функцией премоторной коры, вероятно, служит двустороннее поддержание позы, например фиксация плечей при работе двумя руками и стабилизация бедер при ходьбе. Премоторная кора может вносить вклад в восстановление функции при истинной моторной гемиплегии, возникающей вследствие сосудистого поражения, затрагивающего КСП в лучистом венце. При данном поражении во время ПЭТ-исследования наблюдают повышение активности премоторной коры; кпереди от КСП спускается корково-ретикуло-спинномозговой проводящий путь.
в) Дополнительная моторная зона (ДМЗ). В отличие от премоторной коры, реагирующей на внешние раздражители, ДМЗ (поле 6 на медиальной поверхности полушария) отвечает на внутренние раздражители и принимает особое участие в планировании движений. Это подтверждает тот факт, что ДМЗ возбуждается лобной долей (ДЛПФК) в тот момент, когда мы собираемся совершить движение, даже если движение в итоге не возникает. Важнейшей функцией ДМЗ, вероятно, является предварительное программирование последовательностей движений, уже заложенных в двигательной памяти. ДМЗ функционирует совместно с моторной петлей, проходящей через базальные ганглии, а также взаимодействует с полем 4 и отдает волокна непосредственно в КСП.
Одностороннее поражение ДМЗ может сопровождаться акинезией (невозможностью начать движение) руки и ноги с разных сторон. Двустороннее поражение приводит к тотальной акинезии, включая речевую акинезию.
1. Корковые глазодвигательные поля. На рисунке ниже показаны корковые глазодвигательные поля, участвующие в сканирующих движениях (саккадах). Их взаимодействия и функции представлены в таблице ниже.
2. Дорсолатеральная префронтальная кора (ДЛПФК). ДЛПФК — высший когнитивный центр, участвует в оценке зрительной картины, принятии решения о сознательном совершении саккад и добровольном подавлении рефлекторных саккад (произвольные саккады возникают по воле человека). Рефлекторные саккады — автоматические реакции на объекты, появляющиеся в периферическом поле зрения. Строго говоря, рефлекторные саккады следует называть ответными; они не являются истинными рефлексами, так как способны подавляться волей.
3. Поясная кора. Вместе с ДЛПФК участвует в принятии решений и оценке эмоциональной важности, или ценности, видимых объектов.
4. Дополнительное глазодвигательное поле. Занимает передний отдел ДМЗ и участвует в планировании движений, особенно, если требуются множественные саккадические движения.

5. Фронтальное глазодвигательное поле. Фронтальное глазодвигательное поле (ФГДП) инициирует произвольные саккады, благодаря которым происходит переключение внимания на раздражитель или подавляется стремление к прямому взору на новый раздражитель в ответ на одно или более из трех перечисленных афферентных влияний. ФГДП «поддерживает» карту зрительного пространства относительно глазодвигательных координат и вместе с верхним холмиком играет ключевую роль в создании зрительно-направленных и произвольных саккад; поражения обеих структур приводят к необратимой утрате саккадических движений. Клинические и экспериментальные наблюдения у обезьян показали следующее.
• ФГДП имеют тоническую двустороннюю активность.
• Повышение активности среднего отдела ФГДП с одной стороны приводит к образованию горизонтальных саккад, направленных в сторону противоположного зрительного поля (противоположной саккады).
• Повышение активности верхнего отдела с одной стороны приводит к созданию нисходящей косонаправленной саккады; двусторонняя активация верхнего отдела приводит к обращению взора обоих глаз вертикально вниз.
• Повышение активности в нижнем отделе приводит к аналогичным последствиям относительно взора вверх.
6. Теменное глазодвигательное поле. Теменное глазодвигательное поле (ТГДП) отвечает за создание рефлекторных саккад и побуждает ФГДП к созданию произвольных саккад. ТГДП также участвует в пространственном восприятии, создавая схему зрительной картины.
Поражения префронтальной коры и лобной доли описаны в отдельной статье на сайте.
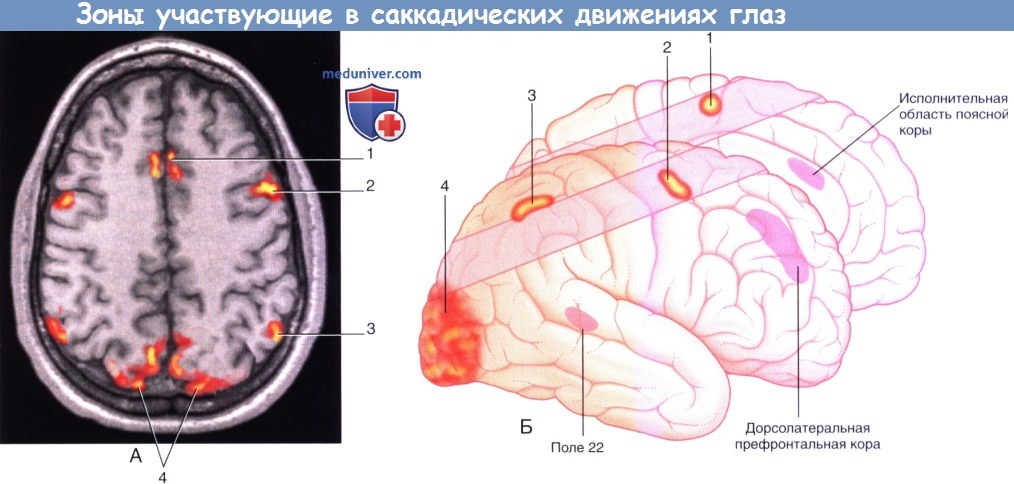 Зоны коры больших полушарий, участвующие в саккадических движениях глаз.
Зоны коры больших полушарий, участвующие в саккадических движениях глаз.
(1) Дополнительное глазодвигательное поле.
(2) Фронтальное глазодвигательное поле.
(3) Теменное глазодвигательное поле.
(4) Ассоциативная зрительная кора.
в) Синдром ригидного человека (СРЧ). Редкое, но легко распознаваемое заболевание, известное как синдром ригидного человека (СРЧ), или синдром мышечной скованности, — аутоиммунное заболевание центральной нервной системы, связанное с наличием циркулирующих антител к глутаматдекарбоксилазе (GAD65),— ключевому ферменту, преобразующему глутамат в ГАМК. Синдром ригидного человека (СРЧ) проявляется скованностью мышц с эпизодическими мышечными спазмами (вызванными одновременным сокращением мышц синергистов и антагонистов, преимущественно в проксимальных отделах конечностей и осевой мускулатуре) и боязнью выполнения определенных задач. В норме контроль верхних моторных нейронов происходит за счет тонической активности прилежащих тормозных ГАМК-ергических вставочных нейронов.
Одни зоны коры поражаются сильнее других, а клинические проявления связаны с нарушением функции этих ГАМК-ергических нейронов, что приводит к гипервозбудимости коры. В настоящее время уточняют роль циркулирующих антител в патогенезе синдрома ригидного человека (СРЧ).
г) Резюме. Кора больших полушарий имеет одновременно пластинчатую и колончатую организацию. К двум основным типам клеток относят пирамидные и непирамидные клетки (вставочные нейроны). Пирамидные клетки расположены в слоях (пластинках) II, III, а также V (в виде веретеновидных клеток) и VI. Слой IV богат шиповатыми звездчатыми клетками (модифицированными пирамидными нейронами). Мелкие пирамидные клетки связывают извилины одного полушария, средние пирамидные клетки связывают между собой соответствующие зоны двух полушарий, а от крупных пирамидных клеток отходят волокна к таламусу, стволу мозга и спинному мозгу. Все корковые волокна считают возбуждающими; шиповатые звездчатые клетки также считают возбуждающими пирамидными клетками.
Корковые вставочные нейроны—тормозные. Колончатая организация представлена колоннами клеток, которые рассматривают в качестве первичной функциональной единицы обработки информации в коре; они состоят из определенных нейронных «микроцепей».
Части тела в соматосенсорной коре представлены инвертированными. Важная входящая информация поступает от вентрального заднего ядра таламуса, а направляется к первичной моторной коре и нижней теменной коре. В первичную зрительную кору входит коленчато-шпорный путь. Клеточные реакции различной сложности зависят от передачи информации с более простых типов клеток на более сложные. Свойство ассоциативных зрительных зон—распознавание черт, например цвета, формы и движения. Обработка цвета и формы продолжается в коре нижележащей височной доли, а движений—в задней теменной доле. Первичная слуховая кора занимает верхнюю поверхность верхней височной извилины, а ассоциативная слуховая кора расположена с латеральной стороны от нее.
Первичная моторная кора занимает прецентральную извилину. Она дает начало большей части волокон пирамидного тракта, а части тела в ней представлены инвертированными. Основная информация подходит к ней от соматосенсорной коры, мозжечка (через вентральное заднее ядро таламуса), премоторной и дополнительной моторной зон. Премоторная кора отвечает преимущественно на внешние раздражители, а дополнительная моторная зона — на внутренние стимулы. Дорсолатеральная префронтальная кора контролирует четыре различных корковых зоны, в различной степени участвующих в создании противоположных саккад.
Редактор: Искандер Милевски. Дата публикации: 22.11.2018